Материал: Черешнев ВА, Шилов, Черешнева. Экспериментальные модели в патологии
|
|
|
|
В-кл |
|
|
Антиген |
|
IL-2, IL-4, IL-5, IL-21, IL-6, |
IFN- , TGF- |
В-клВ-кл Плазматическая |
||
|
|
|
|
|
|
клетка |
|
|
|
CD4CD4 |
|
|
|
|
Tn |
CD4 |
|
|
|
|
|
CD4 |
|
CD4 |
|
|
|
|
|
|
|
|
|
|
DC(АПК) |
|
TfH |
CD4 |
|
|
Синтез |
Наивные |
|
|
|
|||
|
Тh2, |
|
|
В-кл |
антител |
|
|
Тh |
Th1, |
|
|
IgM |
|
|
|
|
В-кл |
В-кл |
||
MHC II |
|
Th3 |
|
|
IgG |
|
|
|
|
|
|
||
+пептид |
|
|
|
В-клетки памяти |
IgA |
|
|
|
|
|
|
||
РАСПОЗНА- |
АКТИВАЦИЯ |
КЛОНАЛЬНАЯ |
ДИФФЕРЕНЦИ- |
ЭФФЕКТОРНАЯ |
||
ВАНИЕ |
АКТИВАЦИЯ |
ЭКСПАНСИЯ |
РОВКА, ПЕРЕКЛЮФУНКЦИЯ |
|||
АНТИГЕНА |
|
ЧЕНИЕ КЛАССА |
|
|||
|
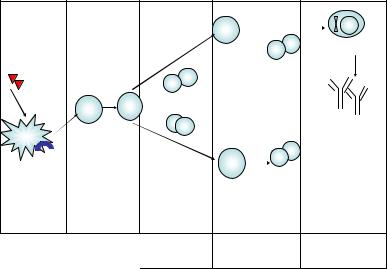
Рис. 15. Развитие первичного иммунного ответа |
|||||
|
с продукцией антител классов IgM, IgG, IgA |
|||||
щийся Tfh-клетками. В эффекторных функциях образовавшихся анти тел, направленных на уничтожение прежде всего внеклеточных пато генов, участвуют клеточные и гуморальные системы врожденного им мунитета. Из них наиболее важную роль играют фагоцитирующие клетки и белки системы комплемента.
Второй вариант ответа направлен на защиту от другой разно видности внеклеточных патогенов – макропаразитов (гельминтов), однако при этом защита обусловлена в наибольшей степени микрофа гами – эозинофилами, которые привлекаются цитокинами IL-3, 5, 13, секретируемыми Th2-лимфоцитами, тучными клетками и базофилами. При этом эозинофилы сорбируются на поверхности паразита, выделя ют из своих эозинофильных гранул высокоактивные белки, которые убивают гельминт, вызывают его деградацию, а макрофаги завершают процесс. Помимо этого в эффекторные реакции вовлечены тучные клетки и базофилы, выброс медиаторов которых запускает локальный воспалительный ответ. В условиях патологии при системном выбросе провоспалительных медиаторов тучными клетками развиваются ал лергические реакции I типа (см. гл. XIII). Переключение синтеза анти
135
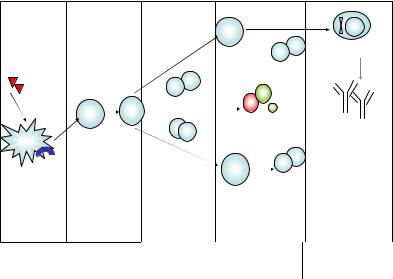
тел на классы IgE и IgG4 происходит на уровне В-лимфоцитов глав |
||||||
ным образом при участии IL-4 и IL-13. Продуцируемый Th2-клетками |
||||||
IL-5 способен непосредственно активировать эффекторные функции |
||||||
эозинофилов без участия антител класса IgE (рис. 16). |
|
|||||
|
|
|
IL-4, IL-5, IL-13 |
В-кл |
|
|
|
|
|
|
В-клВ-кл |
|
|
Антиген |
|
|
|
|
Плазматическая |
|
|
|
|
|
|
|
клетка |
|
IL-4 |
|
CD4CD4 |
|
|
|
|
Tn |
CD4 |
IL-3, IL-5 |
|
|
|
|
|
Эозинофилы, |
|
|||
|
CD4 |
|
CD4 |
|
||
|
|
|
тучные клетки, |
|
||
DC(АПК) |
|
Тh2 |
CD4 |
Синтез |
||
Наивные |
|
базофилы |
|
|||
|
Тh |
|
|
|
В-кл |
антител |
MHC II |
|
|
|
В-кл |
В-кл |
IgE и |
|
|
|
|
IgG4 |
||
+пептид |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
В-клетки памяти |
|
|
РАСПОЗНА- |
АКТИВАЦИЯ |
КЛОНАЛЬНАЯ |
ДИФФЕРЕНЦИ- - |
ЭФФЕКТОРНА |
||
ЭФФЕКТОРНАЯ |
||||||
ВАНИЕ |
АКТИВАЦИЯ |
ЭКСПАНСИЯ |
РОВКА |
|
ФУНКЦИЯ |
|
АНТИГЕНА |
|
ЦИРОВКА |
Я ФУНКЦИЯ |
|||
Рис. 16. Развитие первичного иммунного ответа по Th2-типу |
||||||
Третий вариант гуморального иммунного ответа развивается на тимуснезависимые антигены без участия Т-хелперов. К тимуснеза висимым антигенам относятся бактериальные гетерополисахариды, харатеризующиеся жесткой структурой и наличием часто повторяю щихся антигенных детерминант (например, пневмококковый полиса харид); бактериальные липополисахариды, обладающие адъювантным действием (например, липополисахарид E. coli); некоторые синтетиче ские антигены. В ответе на эти антигены участвуют минорные субпо пуляции В-клеток: В1-лимфоциты и В-лимфоциты маргинальной зоны селезенки.
Th17-тип иммунного ответа. Th17-клетки – субпопуляция Т хелперов, открытая несколько лет назад. Свое название эти клетки получили в связи с тем, что они продуцируют IL-17A, E (или IL-25) и F, относящиеся к семейству IL-17 (которое помимо названных цито кинов включает IL-17B, C и D). Помимо этого Th17-клетки продуци
136
руют IL-21 и IL-22. Th17-тип иммунного ответа имеет сходство как с Th1-, так и с Th2-ответами. Th17-клетки обеспечивают резистентность к возбудителям Listeria, Salmonella, Toxoplasma, Cryptococcus, Leish mania и Francisella. Преимущественная продукция IL-17 отмечается при инфекциях, вызванных Borrelia burgdoferi, Mycobacterium tuberculosis, Bacteroides fragilis, грибами. Существенную и специфич ную роль Th17-ответ имеет в таких экспериментальных моделях у грызунов, как поражение легких Klebsiella, инфекция, развивающаяся при внутривенном введении Candida albicans, а также вызванная в кишечнике естественным патогеном Citrobacter rodentium. Кроме уча стия в контроле инфекций Th17-клетки играют важную роль в индук ции и прогрессировании аутоиммунных заболеваний, в частности рас сеянного склероза. В качестве вторичных эффекторных клеток в реак ции Th17-типа вовлекаются гранулоциты и клетки моноцитарно макрофагального ряда.
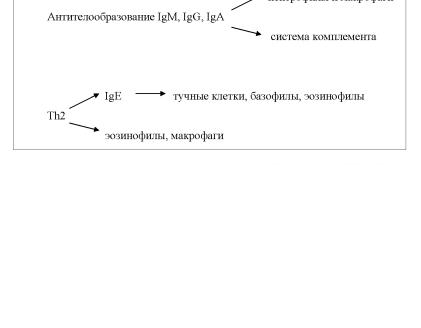
Таким образом, в динамике адаптивного иммунного ответа помимо антител как эффекторных молекул и цитотоксических Т лимфоцитов в элиминацию антигенов и клеток, несущих их, вовлека ются клеточные и гуморальные системы врожденного иммунитета. Главными клеточными эффекторами при реакциях Th1-типа выступа ют макрофаги и NK-клетки, при антителообразовании с продукцией IgM, IgG, IgA – нейтрофилы и макрофаги, при реакциях Th2-типа – эозинофилы, тучные клетки, базофилы и макрофаги. Основной гумо ральной эффекторной системой врожденного иммунитета является система комплемента (рис. 17).
При сопоставлении эффекторных механизмом врожденного и адаптивного иммунитета нетрудно заметить, что в элиминации пато гена используются почти идентичные механизмы. Эффекторные анти генспецифические молекулы и клетки адаптивного иммунитета при дают более высокую специфичность эффекторным механизмам врож денного иммунитета и усиливают их за счет контактных межклеточ ных взаимодействий и стимулирующего действия цитокинов. Во всех вариантах основной эффекторной клеткой, завершающей элиминацию патогена, является макрофаг.
Накопление иммунокомпетентных клеток и концентрация антител в очаге воспаления
Основной территорией, на которой происходит активация, пролиферация, дифференцировка клеток при развитии адаптивного иммунного ответа с образованием антител и/или эффекторных субпо пуляций Т-лимфоцитов, являются тимусзависимые и тимуснезависи-
137
по сути, мостик между врождённым и приобретенным иммунитетом. Для поступления в очаг воспаления антител, а также гуморальных эф фекторов врожденного иммунитета важное значение имеет повышение проницаемости капилляров и увеличение гидростатического давления, связанное с развитием венозной гиперемии.
Деструкция иммунокомпетентными клетками и эффекторными молекулами антигенов и клеток, поврежденных патогеном
Полное разрушение патогена в очаге воспаления происходит в результате реализации эффекторных механизмов, связанных с актива цией белков системы комплемента и комплемент-зависимым цитоли зом, опсонизацией при фагоцитозе, деструкцией фагоцитами при уча стии радикалов кислорода и оксида азота, NK-клетками и цитотокси ческими Т-лимфоцитами клеток, несущих антигены, PAMP или
DAMP.
Выведение продуктов распада общеорганизменными системами выделения
Низкомолекулярные метаболиты, образовавшиеся в предше ствующей стадии удаляются из организма через системы выделения – почки и желудочно-кишечный тракт.
Регенерация. Выздоровление. Поствоспалительная иммуносупрессия с сохранением иммунологической памяти
В организме, санированном от патогена, развивается послед няя, завершающая стадия – продолжается регенерация, наступает вы здоровление, отмечается спад продуктивного иммунного ответа с по явлением иммунологической памяти.
Иммунологическая память – это приобретенная способность иммунной системы отвечать более быстро и эффективно на повторный контакт с тем же антигеном в виде антигенспецифических гумораль ных и клеточных реакций.
Благодаря клеткам памяти, которые длительное время перси стируют в организме (часто на протяжении всей жизни), при повтор ном попадании того же антигена в организм вторичный иммунный ответ развивается значительно быстрее (сокращается продолжитель ность индуктивного периода) и интенсивнее, причем продукция анти тел детерминирована строго генетически. В ранние сроки первичного иммунного ответа до 80% антител приходится на IgM, а вторичного – до 90% приходится на IgG. При вторичном иммунном ответе нет ха рактерного для первичного последовательного переключения с IgM на
139