Материал: Черешнев ВА, Шилов, Черешнева. Экспериментальные модели в патологии
лимфоцитов обязательно находится клетка с соответствующим рецеп тором, комплементарным данному антигену. Этот механизм объясня ется клонально-селекционной теорией Ф. Бернета (F. Burnet). Проис ходит селекция для дальнейшей активации соответствующих клонов лимфоцитов и развивается адаптивный иммунный ответ.
Развитие адаптивного иммунного ответа
Дальнейшие события прежде всего определяются особенно стями поступающего в организм антигена. Антиген захватывается ден дритной клеткой, процессируется и в виде комплекса МНС (I или II классов)+пептид представляется соответствующему (специфическому) клону Т-лимфоцитов. Зрелые Т-лимфоциты помимо Т-клеточного ан тигенраспознающего рецепторного комплекса несут на своей мембра не корецепторные молекулы CD4 (Т-хелперы, корецептор к MHC II класса) или CD8 (цитотоксические Т-лимфоциты и их предшественни ки, корецептор к MHC I класса).
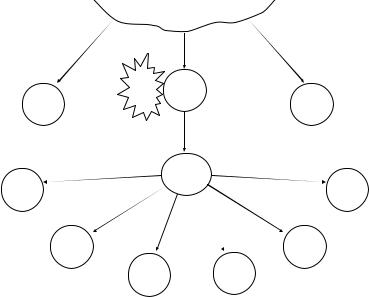
После распознавания лиганда (антигенный пептид + MHC II класса) наивные Т-хелперы (Тh) начинают дифференцироваться в суб популяции зрелых Th1-, Th2-, Th9-, Th17-, Tfh-клеток, т.е. активаци онную эстафету от дендритных клеток принимают Т-лимфоциты хелперы (рис. 13).
Каждый из этих типов лимфоцитов отвечает за свой комплекс реакций, особенно полно он изучен для Th1 и Th2, в меньшем объеме
– для Th9, Th17, Tfh. Характеристика этих клеток и их функции пред ставлены в табл. 7.
Недавно открытые Th9-клетки обеспечивают противопарази тарный иммунитет, Th17 играют центральную роль в аутоиммунных процессах, Tfh обеспечивают помощь активированным В-лимфоцитам в лимфоидных фолликулах периферических органов иммунной систе мы (селезенка, лимфатические узлы, лимфоидные образования слизи стых). Все варианты иммунного ответа контролируются регуляторны ми Т-лимфоцитами (Тreg), имеющими фенотип CD4+CD25+Foxp3+. Выделяют две разновидности этих клеток – естественные nTreg (регу лируют другие клетки без предварительной активации антигеном) и индуцибельные iTreg (активируются антигеном). Эти клетки продуци руют тормозные цитокины IL-10 и TGF-β и являются функциональ ным аналогом ранее описанных супрессорных лимфоцитов.
В зависимости от преимущественной активации тех или иных субпопуляций Th, которая во многом определяется цитокиновым мик роокружением, возможна поляризация иммунного ответа с преимуще ственной активацией разных типов вторичных эффекторных клеток.
130
ТИМУС
АПК |
Наивные |
|
nTreg |
Th |
NKT |
|
Активи- рованные
iTreg |
Th |
Th17 |
|
Tfh |
Th1 |
|
|
|
|
Th9 |
Th2 |
Рис. 13. Схема развития субпопуляций Т-хелперов после распознавания антигенных пептидов
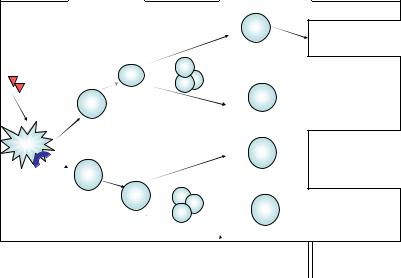
Th1-тип иммунного ответа. В первом варианте Th1-ответа (рис. 14) активированные после повторного контакта с антигенными пептидами в комплексе с MHC II класса (на поверхности макрофага или другой антигенпрезентирующей клетки) Th1-клетки реализуют свою активность путем взаимодействия с макрофагами, выступающи ми в качестве вторичных эффекторных клеток при реакции ГЗТ. Th1 клетки активируют макрофаги, передавая костимулирующий сигнал через взаимодействие CD154 (на поверхности Th1-клетки) c молеку лой CD40 (на мембране макрофага), а также через секретируемый ими цитокин IFN-γ. Этот двунаправленный диалог приводит к мощной до полнительной активации макрофагов, усилению всех факторов микро бицидности и формированию нового микробицидного агента – оксида азота. Благодаря экспрессии макрофагами индуцибельной NO-синтазы фагоцитоз становится завершенным. Помимо клеток моноцитарно макрофагального ряда активированные Th1-клетки в качестве вторич ных эффекторов используют и NK-клетки.
131

|
|
Субпопуляции CD4+ Т-хелперов, их дифференцировка и функции |
|
Таблица 7 |
||||||||
|
|
|
|
|||||||||
Свойства |
|
Th1 |
|
Th2 |
|
Th9 |
|
Th17 |
|
Tfh |
|
Treg |
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
Продукция |
|
IFN- , IL-2, TNF- , |
|
IL-4, IL-5, |
|
IL-9 |
|
IL-17A, |
|
IL-21 |
|
TGF-β, IL-10 |
цитокинов |
|
TNF- |
|
IL-10, IL-13 |
|
|
|
IL-17F |
|
|
|
|
|
|
|
|
|
|
|
|
IL-21, IL-22, IL-25, |
|
|
|
|
|
|
|
|
|
|
|
|
IL-26, TNF- |
|
|
|
|
Транскрипционные |
|
STAT-4, |
|
STAT6, GATA3, |
|
GATA3, Smads, |
|
ROR- , ROR- t |
|
BCL6, MAF |
|
Foxp3 |
факторы |
|
T-bet, Hlx |
|
c-maf, IRF4, Gfi-1 |
|
STAT6 |
|
|
|
|
|
|
Индуцирующие |
|
IL-12, IL-18, IL-27, |
|
IL-4 |
|
TGF-β, |
|
IL-6, TGF-β, |
|
IL-21, IL-6 |
|
TGF-β, IL-10, |
цитокины |
|
IFN- |
|
|
|
IL-4 |
|
IL-1 и/илиIL-1 , |
|
|
|
IL-2, TSLP |
|
|
|
|
|
|
|
|
IL-21, IL-23 |
|
|
|
|
Патогены |
|
Вирусы, |
|
Внеклеточные |
|
Гельминты |
|
См. |
|
Внеклеточные |
|
Негативная |
|
|
внутриклеточные |
|
патогены, втом |
|
|
|
текст |
|
патогены, |
|
регуляцияклиренса |
|
|
бактерии, |
|
числегельминты, |
|
|
|
|
|
доступныедля |
|
патогенов |
|
|
простейшие, |
|
доступныедля |
|
|
|
|
|
антителклассов |
|
|
|
|
грибы |
|
антителклассов |
|
|
|
|
|
IgM, IgG, IgA |
|
|
|
|
|
|
IgE иIgG4 |
|
|
|
|
|
|
|
|
Иммунопатология |
|
Аутоиммунные |
|
Аллергические |
|
|
|
Аутоиммунные |
|
|
|
Поддержание |
|
|
заболевания, |
|
реакции Iтипа |
|
|
|
заболевания, |
|
|
|
иммунологической |
|
|
реакция |
|
|
|
|
|
аллергия |
|
|
|
толерантности |
|
|
трансплантат |
|
|
|
|
|
|
|
|
|
|
|
|
против хозяина |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
IFNγ, IL-2 |
|
|
|
|
CD4 |
Активация |
|
|
|
|
макрофагов, NK,- |
Антиген |
|
|
Т-клетки |
В-клеток и др. |
|
|
|
||
CD4 |
CD4 |
эффекторы |
|
|
|
|
|
||
|
CD4 |
|
|
|
|
|
CD4 |
|
|
|
Th |
|
CD4 |
|
|
CD4 |
|
Т-клетки памяти |
|
|
|
|
|
|
DC(АПК) |
|
|
|
Киллинг |
|
|
CD8 |
инфицированных |
|
|
|
|
||
|
|
|
клеток-мишеней |
|
|
Th |
|
|
|
MHC+ |
|
Т-клетки эффек- |
(перфорин- |
|
пептид |
CD8 |
|
торы (ЦТЛ) |
гранзимовый) |
CD8 |
CD8 |
|
||
|
|
|
||
|
|
CD8 |
CD8 |
|
|
|
CD8 |
|
|
|
|
|
Т-клетки памяти |
|
РАСПОЗНА- |
АКТИВАЦИЯ |
КЛОНАЛЬНАЯ |
ДИФФЕРЕНЦИ- - |
ЭФФЕКТОРНАЯ |
ВАНИЕ |
АКТИВАЦИЯ |
ЭКСПАНСИЯ |
РОВКА |
ФУНКЦИЯ |
АНТИГЕНА |
|
ЦИРОВКА |
Я ФУНКЦИЯ |
Рис. 14. Развитие первичного иммунного ответа по Th1-типу
Второй вариант Th1-ответа (см. рис. 14) реализуется через цитотоксические Т-лимфоциты (ЦТЛ). Эти клетки предназначены для защиты от вирусов и патогенов, живущих в цитозоле. Зрелые актив ные ЦТЛ и их наивные предшественники несут на своей мембране корецептор CD8, что позволяет распознавать «чужие» антигенные пептиды в комплексе с MHC I, которые представлены на мембране всех ядросодержащих клеток организма (в отличие от MHC II, кото рые находятся на мембране только профессиональных антигенпрезен тирующих клеток). Эта особенность позволяет ЦТЛ осуществлять эф фективный надзор за вирусными инфекциями, опухолевой трансфор мацией, мутациями – процессами, которым могут быть подвержены любые ядросодержащие клетки. Помощь от Th1-клеток состоит глав ным образом в снабжении цитотоксических Т-лимфоцитов ростовым цитокином IL-2, необходимым для достаточного накопления этих кле ток. ЦТЛ выполняют функцию киллеров: уничтожают инфицирован ные клетки вместе с патогеном, а также опухолевые клетки, участвуют в отторжении трансплантатов. При этом используется перфорин гранзимовый механизм: из гранул ЦТЛ на мембрану клетки-мишени сначала действуют перфорины, в результате чего образуются поры
133
диаметром 16 нм, через которые из тех же гранул внутрь клетки про сачиваются гранзимы, которые инициируют программу апоптоза. При этом разрушается и сама инфицированная клетка, и вирусы в ней. Возможна и контактная форма запуска апоптоза клетки-мишени без цитолиза за счет взаимодействия находящегося на мембране ЦТЛ FasL с мембранным рецептором апоптоза клетки-мишени CD95 (Fas). Ос татки погибших клеток утилизируются макрофагами.
Антителообразование, или гуморальный иммунный ответ,
реализуется в трех вариантах:
1)тимусзависимый иммунный ответ с последовательным пе реключением при участии всех типов Th-клеток класса образующихся антител с IgM на IgG и на IgA;
2)тимусзависимый Th2-тип иммунного ответа с переключе нием класса синтезируемых антител на IgE и IgG4;
3)тимуснезависимый иммунный ответ с продукцией только IgM антител.
Первый вариант ответа развивается на большинство тимусза висимых антигенов. Для активации В-лимфоцитов помимо антиген специфического сигнала с антигенраспознающих В-клеточных рецеп торов (распознающая часть рецептора у наивных В-лимфоцитов – мембранные IgM и IgG, они способны в отличие от Т-клеток взаимо действовать с антигеном в нативной форме) необходимы сигналы со стороны Т-хелперов, реализуемые как за счет межклеточного контак та, так и короткодистантно при участии цитокинов. Контактные взаи модействия носят двунаправленный характер. С одной стороны, В клетка сама выступает в роли антигенпрезентирующей клетки: погло тив антиген, обрабатывает его, встраивая антигенный пептид в состав молекулы MHC II-класса, презентирует этот комплекс T-хелперу. С другой стороны, она получает активирующий сигнал от Т-хелпера за счет взаимодействия CD40 (на мембране В-лимфоцита) с CD154 (на мембране Т-лимфоцита). В ходе контактных взаимодействий происхо дит максимальное сближение клеток при участии молекул клеточной адгезии на расстояние 15 нм с формированием иммунологического синапса. В короткодистантных взаимодействиях последовательно уча ствуют цитокины многих субпопуляций T-хелперов (рис. 15). Так, для образования из В-лимфоцитов плазматических клеток, продуцирую щих антитела класса IgM, необходимы IL-2 (Th1-клетки), а также IL-4
иIL-5 (Th2-клетки); переключения на IgG – IFN- (Th1-клетки), а на IgA – TGF- (nTreg-, iTreg-клетки). Ключевую роль в индукции про лиферации, дифференцировки в направлении плазматических клеток и переключения классов иммуноглобулинов играет IL-21, продуцирую-
134