Материал: Normalnaya_fiziologia_ekzamen2021
Строение и классификация синапсов. Механизмы синаптической передачи.
Синапсы – специализированная структура, которая обеспечивает передачу нервного импульса из нервного волокна на эффекторную клетку – мышечное волокно, нейрон или секреторную клетку.
Синапсы – места соединения нервного отростка (аксона) одного нейрона с телом или отростком (дендритом, аксоном) другой нервной клетки (прерывистый контакт между нервными клетками).
Все структуры, обеспечивающие передачу сигнала с одной нервной структуры на другую – синапсы.
Значение – передает нервные импульсы с одного нейрона на другой, обеспечивает передачу возбуждения по нервному волокну (распространение сигнала).
Большое количество синапсов обеспечивает большую площадь для передачи информации.
Виды синапсов по расположению:
1)аксодендритические синапсы - на дендритах и теле нейронов. Передатчики – аксоны;
2)аксосоматические синапсы - между аксоном и телом нейрона;
3)аксошипиковые синапсы - на шипиках (выросты на дендритах. С их изменением меняется работа нейронов);
4)аксоаксональные синапсы - между аксонами нейронов;
5)дендродендритические синапсы - между дендритами нейронов;
6)сомосоматические синапсы - между телами нейронов.
Виды синапсов по способу передачи сигналов:
1)химические синапсы – возбуждение передается посредством медиаторов;
2)электрические синапсы - возбуждение передается посредством ионов;
3)смешанные синапсы - возбуждение передается посредством и медиаторов, и ионов.
Виды синапсов по анатомо-гистологическому принципу:
1)нейросекреторные;
2)нервно-мышечные;
3)межнейронные.
Виды синапсов по нейрохимическому принципу:
1)адренергические – медиатор норадреналин;
2)холинэргические – медиатор ацетилхолин.
Виды синапсов по функциональному принципу: возбуждающие и тормозные.
Между окончаниями двигательного нейрона и мышечным волокном существует нервно-мышечное соединение, отличающееся по строению, но сходное в функциональном отношении синаптическими контактами.
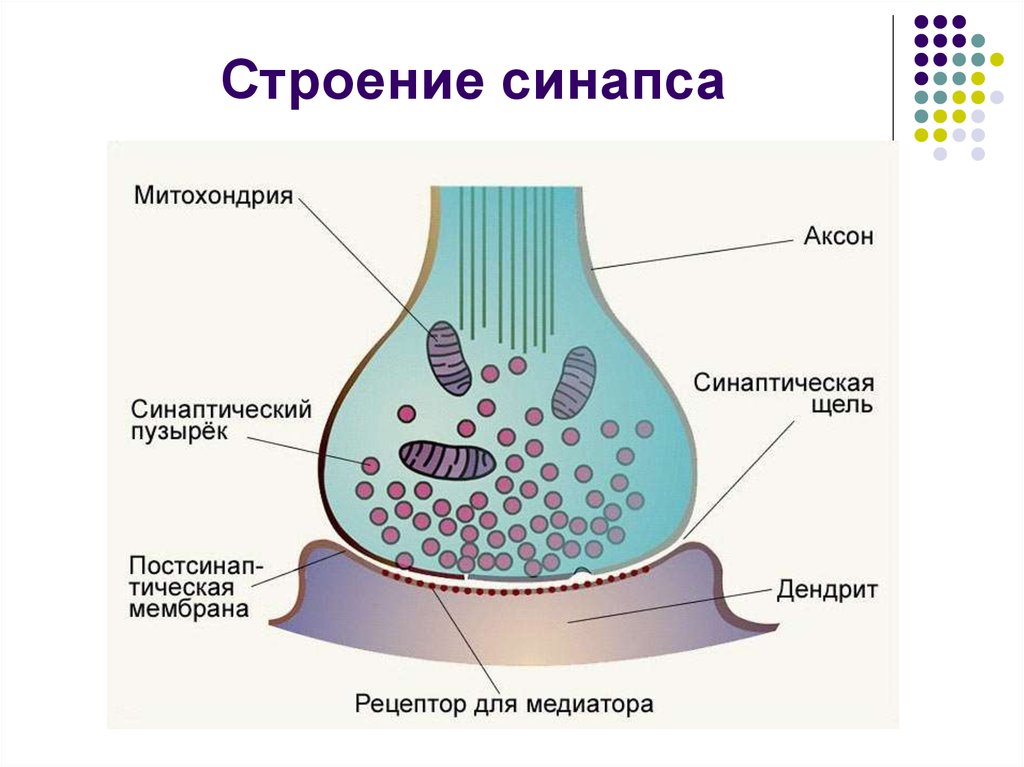
Строение синапса:
1)пресинаптическая мембрана - принадлежит нейрону, от которого передается сигнал;
2)синаптическая щель, заполненная жидкостью с высоким содержанием ионов Са;
3)постсинаптическая мембрана - принадлежит клеткам, на которые передается сигнал.
Между нейронами всегда существует перерыв, заполненный межтканевой жидкостью.
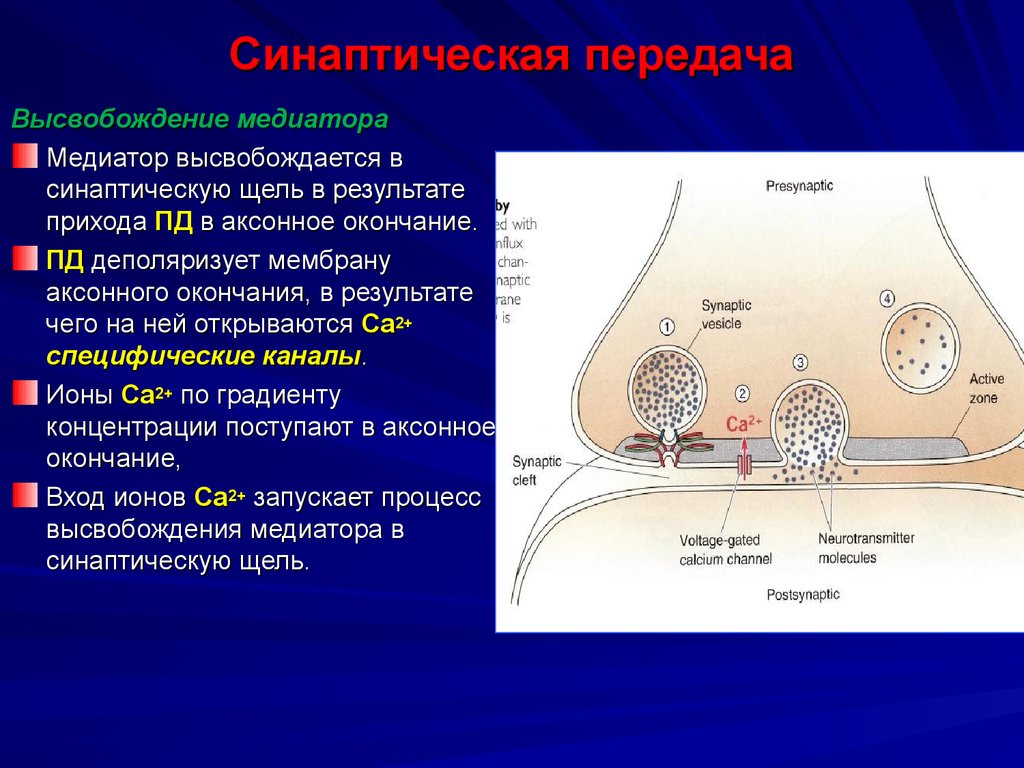
Когда возбуждение приходит в область пресинаптической мембраны, она деполяризуется, это активирует кальциевые потенциалзависимые каналы N-типа, они медленные, быстро инактивируются. В клетку входит небольшое количество кальция. Вошедший кальций связывается со специальным белком (кальций - связывающий белок) - синаптосин. Кальций активирует фосфолирирование нескольких молекул синаптосина, вызывая их конформацию, что приводит к ослаблению связей везикул с цитоскелетом клетки и они (везикулы) перемещаются к внутренней поверхности пресинаптической мембраны и прилипает к ней. Это вызывает разрыв везикул (ферментативный протеолиз). Параллельно прилипание к мембране активирует мембранный белок-синаптопор, который выступает как переносчик медиатора через мембрану либо как белок-переносчик за счет конформации молекулы, либо образуя транспортный канал, либо вызывая экзоцитоз.
Медиатор диффундирует в синаптическую щель, он не выходит за пределы синаптической щели, т.к. сбоку имеются синаптические барьеры, которые препятствуют выходу медиатора за пределы синапса.
Молекулы медиатора идут к постсинаптической мембране, в область субсинаптической мембраны, которая имеет много однотипных хеморецепторов и образуют комплекс «медиатор – рецептор». Это вызывает активацию соответствующих рецепторуправляемых ионных каналов.
Все медиаторы можно разделить на возбуждающие медиаторы и тормозные медиаторы.
- Возбуждающие медиаторы взаимодействуя с рецептором субсинаптической мембраны вызывают активацию натриевых каналов и формируют входящий натриевый ток, который вызывает возникновение частичной деполяризации, т.е. рецепторный потенциал, который на уровне синапс обозначают как возбуждающий постсинаптический потенциал.
- Тормозной медиатор вызывает усиление входящего калиевого тока или входящего ток хлора, т.е. вызывает локальную гиперполяризацию. Это формирует тормозной постсинаптический потенциал. Конечный эффект (потенциал действия или тормозной потенциал) формируется за счет суммации.
В синапсе известно два вида суммации:
1)пространственная суммация локальных очагов возбуждения - в пространстве суммируются эти локальные очаги и возникает потенциал действия;
2)временная суммация - на одни и те же рецепторы последовательно через короткие интервалы времени воздействуют новые порции медиатора и возникает как бы ступенька - частичная деполяризация, она не окончилась, на нее наслаивается следующая частичная деполяризация и так поляризация достигает КУДа (критический уровень деполяризации).
В дальнейшем комплекс «медиатор-рецептор» диссоциирует. Если этого не происходит, то в возбуждающих синапсах возникает стойкая деполяризация по типу катодической депрессии. При этом данный рецептор перестает воспринимать какую-либо другую информацию. Поэтому в нормальных, естественных условиях медиатор отсоединяется от рецепторов и разрушается ферментами (холинэстераза и т.д.), которые имеются в синапсе. Другой способ инактивации медиатора – аптейк- обратный захват пресинаптической мембраной. За счет этого синапс экономно расходует медиатор.
Характерные признаки процесса синаптической передачи:
1)односторонний характер проведения возбуждения в синапсе от пре- к постсинаптической мембране;
2)квантовый (парциальный) характер освобождения медиатора;
3)количество квантов медиатора пропорционально частоте и силе приходящего к синапсу (пресинаптической мембране) нервного раздражения;
4)синаптическая передача не подчиняется закону "все или ничего";
5)синапс способен к суммации процессов возбуждения;
6)проведение возбуждения в синапсе осуществляется с задержкой во времени (синаптическая задержка);
7)при многократном прохождении возбуждения через синапс возникает эффект облегчения проведения возбуждения - возникающее возбуждение наслаивается на остаточные процессы;
8)для синапса характерно проведение возбуждения с декрементом (с ослаблением по силе);
9)трансформация - способность синапса изменять частоту пришедшего раздражения (как правило, синапс резко снижает частоту пришедшего раздражения);
10)лабильность синапса существенно меньше, чем у нервов;
11)из всех звеньев рефлекторной дуги синапс - наиболее утомляемый и наиболее чувствительный к ядам и недостатку кислорода элемент цепи.
Частное свойство: длительное или очень сильное воздействие на синапс приводит к прекращению синаптической передачи, которое обусловлено истощением медиатора в области пресинаптической мембраны (эффект истощения).
Все вышеперечисленные закономерности характерны как для синапсов ЦНС, так и для периферических синапсов.
Нервный центр: определение, строение. Свойство пространственной и временной суммации в нервном центре.
Нервный центр — совокупность структур центральной нервной системы, координированная деятельность которых обеспечивает регуляцию отдельных функций организма или определенный рефлекторный акт.
Основными клеточными элементами нервного центра являются многочисленные нейроны, скопление которых формирует нервные ядра. В состав центра могут входить нейроны, рассеянные за пределами ядер. Нервный центр может быть представлен структурами мозга, располагающимися на нескольких уровнях центральной нервной системы (центры регуляции дыхания, кровообращения, пищеварения).
Любой нервный центр состоит из ядра и периферии.
Ядерная часть нервного центра представляет собой функциональное объединение нейронов, в которое поступает основная информация от афферентных путей. Повреждение этого участка нервного центра приводит к повреждению или существенному нарушению осуществления данной функции.
Периферическая часть нервного центра получает небольшую порцию афферентной информации, и ее повреждение вызывает ограничение или уменьшение объема выполняемой функции.
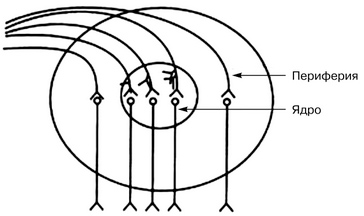
Схема общего строения нервного центра ↑
В нервных центрах выделяют следующие иерархические отделы: рабочие, регуляторные и исполнительные.
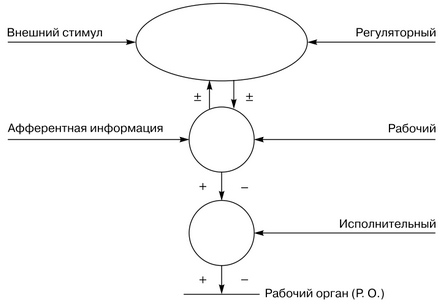
Схема иерархического подчинения разных отделов нервных центров ↑
Рабочий отдел ответствен за осуществление данной функции.
Регуляторный отдел - центр, расположенный в КБПГМ (коре больших полушарий головного мозга) и регулирующий активность рабочего отдела нервного центра. В свою очередь, активность регуляторного отдела нервного центра зависит от состояния рабочего отдела, который получает афферентную информацию, и от внешних стимулов среды.
Исполнительный отдел - двигательный центр, расположенный в спинном мозге и передающий информацию от рабочего отдела нервного центра к рабочим органам.
Временная суммация возникает как в синапсах вследствие того, что чем больше поступает нервных импульсов, тем больше выделяется нейромедиатора в них, тем выше амплитуда возбуждающего постсинаптического потенциала. Поэтому рефлекторная реакция может возникать на несколько последовательных подпороговых раздражений. Пространственная суммация наблюдается тогда, когда к нервному центру идут импульсы от нескольких рецепторных нейронов. При действии на них подпороговых стимулов, возникающие постсинаптические потенциалы суммируются, и в мембране нейрона генерируется распространяющийся потенциал действия.
Общие свойства нервных центров (одностороннее проведение возбуждения, центральная задержка, посттетаническая потенциация, последействие, тонус, трансформация ритма, утомляемость).
Основные свойства нервных центров:
1)одностороннее проведение возбуждения (в нервных центрах прохождение импульсов возбуждения по цепи нейронов осуществляется только в одном направлении: от чувствительного нерва через промежуточные к двигательному, а от двигательного — к органу-исполнителю. Это обусловлено односторонней синаптической передачей импульсов от одной нервной клетки к другой с помощью медиатора, который выделяется концевым аппаратом аксона и содержится лишь в пресинаптической щели. В связи с этим поток нервных импульсов в рефлекторной дуге имеет определенное направление. Это свойство обеспечивает координирующую роль центральной нервной системы и способствует замыкательной функции условно-рефлекторных связей);
2)центральная задержка (замедление проведения возбуждения или латентный (скрытый) периодом рефлекса. Обусловлена более медленным проведением нервных импульсов через синапсы. Интервал от начала раздражения рецептора до появления ответной реакции — 0,2–0,5 с. Чем сложнее рефлекс, тем длительнее центральная задержка. Необходимо учитывать скрытый период при выработке условного рефлекса и подкрепляющий раздражитель надо применять не раньше 0,5 с после воздействия сигнального раздражителя. Скрытый период рефлекса увеличивается при утомлении и заболевании);
3)посттетаническая потенциация - интенсивность ответной реакции нервного центра зависит от предшествующих ритмических раздражений. Явление потенциации – увеличение амплитуды постсинаптического потенциала связано с накоплением ионов кальция в пресинаптическом окончании и вследствие этого при ритмической стимуляции каждый пресинаптический потенциал вызывает высвобождение все большего числа квантов медиатора. Такую же природу имеет феномен посттетанической потенциации: увеличение числа квантов медиатора, высвобождаемых нервным импульсом после предшествующего ритмического раздражения, приводит к повышению возбудимости ПСМ нейронов и всего нервного центра. Может длиться от нескольких минут до нескольких часов в различных структурах мозга. Предполагают, что этот феномен присутствует в пластических перестройках функций синапсов и лежит в основе механизмов организации условных рефлексов и памяти;
4)последействие (присуще всем нервным центрам и характеризуется тем, что на короткое раздражение проявляется длительная ответная реакция, т. е. рефлекторный акт длится еще некоторое время после того, как раздражение рецептора прекращено. Такое последействие объясняется тем, что нервные импульсы от рецепторов поступают к двигательным центрам по различным нервным путям неодновременно: по коротким быстрее, чем по длинным. Запаздывающие импульсы поддерживают возбужденное состояние соответствующего нервного центра. Остаточное возбуждение в нервных центрах может сохраняться до 2 с и способствовать лучшей замыкательной функции при образовании условных рефлексов. После окончания действия любого раздражителя требуется некоторое время для спада остаточного возбуждения нервного центра и освобождения рефлекторных путей для новой рефлекторной деятельности);
5)тонус (нервные центры обладают свойством постоянно находиться в состоянии незначительного возбуждения при относительном рефлекторном покое. Постоянно посылают импульсы, обеспечивающие тоническое сокращение скелетной мускулатуры. Тонус нервных центров поддерживается действием гуморальных веществ и непрерывным потоком импульсов, поступающих от рецепторов. Огромное значение в поддержании мышечного тонуса имеют нервные центры продолговатого, среднего и промежуточного мозга. Тоническое состояние нервных центров и мышечной системы обеспечивает устойчивую выработку условных рефлексов и хорошую работоспособность);
6)трансформация ритма (способность нейронов изменять ритм приходящих импульсов. Механизмами могут быть: возникновение длительного ВПСП (возбуждающий постсинаптический потенциал) на фоне которого возникает несколько спайков; следовые колебания мембранного потенциала. Если следовое колебание достигнет критического уровня деполяризации мембраны, возникнет вторичный спайк; трансформация ритма может быть связана с наличием в нервных цепях реверберационных кругов, повышающих заданный ритм, или тормозных нейронов, понижающих ритм на выходе из нервного центра);
7)утомляемость (проявляется в постепенном снижении и в конечном итоге полным прекращением рефлекторного ответа при продолжительном раздражении афферентных нервных волокон. Связано с нарушением передачи возбуждения в межнейронных синапсах. Нарушение может быть обусловлено резким уменьшением запасов синтезированного медиатора в нервных окончаниях, уменьшением чувствительности к медиатору постсинаптической мембраны нервной клетки, уменьшением ее энергетических ресурсов).
Механизмы взаимодействия нервных центров (принципы координации рефлекторной деятельности): реципрокность, общий конечный путь, субординация, обратная афферентация, доминанта, индукция.
Основные принципы координации:
1)принцип сопряженного торможения или реципрокности (возникновение при возбуждении одного центра одновременного торможения другого центра для обеспечения цепных рефлексов и сложной функции. Пример - регуляция спинным мозгом противоположных по функциональному назначению мышц конечностей. При возбуждении мотонейронов, иннервирующих мышцы сгибатели правой ноги, реципрокно тормозятся мотонейроны мышц разгибателей этой ноги и возбуждаются мотонейроны мышц разгибателей левой ноги. Формирующийся цепной характер рефлексов вслед за этим вызывает возбуждение мотонейронов разгибателей правой ноги и реципрокно — торможение мотонейронов сгибателей правой ноги и возбуждение мотонейронои сгибателей левой ноги. Реципрокные взаимоотношения между указанными рефлексами обеспечивают цепной шагательный рефлекс);
2)принцип общего конечного пути (или принцип конвергенции. Открыт и исследован Ч.С. Шеррингтоном, 1896 г. В нервных центрах количество афферентных (приносящих) клеток намного больше, чем количество эфферентных (выносящих) нейронов, несущих возбуждение к мышцам. Получается, что между нейронами идет борьба "за общий конечный путь", т.е. за то, чтобы передать своё возбуждение на эффентные нейроны. Этот принцип получил также образное наименование "воронки Шеррингтона");
3)принцип субординации (или принцип подчинения, проявляется в виде регулирующего влияния выше расположенных нервных центров на ниже расположенные. Например, двигательные центры головного мозга управляют спинальными мотонейронами. Примером такого влияния является феномен центрального торможения спинальных рефлексов открытый И.М.Сеченовым и получивший название сеченовского торможения. В эксперименте И.М.Сеченова раздражение зрительных бугров лягушки с помощью кристаллика поваренной соли (т.е. раздражение ретикулярной формации среднего мозга) приводило к торможению спинальных двигательных рефлексов, вызываемых погружением лапки лягушки в слабый раствор кислоты. Торможение центров спинного мозга явилось следствием возбуждения центров среднего мозга. Прекращение этого тормозного контроля при перерыве цереброспинальных проводящих путей вызывает резкое повышение возбудимости спинальных центров и гиперрефлексию);
4)принцип обратной афферентации (рецепторное восприятие результатов рефлекторного акта и проведение информации назад в структуры нервного центра, где она обрабатывается и сравнивается с сохраняющимися параметрами возбуждения. Реализуется в виде положительной или отрицательной обратной связи. С помощью обратной афферентации нервные центры осуществляют непрерывный контроль эффективности, целесообразности и оптимальности рефлекторной деятельности);
5)принцип доминанты (был открыт А.А.Ухтомским. Доминанта - общий принцип деятельности нервной системы, проявляющийся в виде господствующей в течение определенного времени системы рефлексов, реализуемых доминирующими центрами, которые подчиняют себе или подавляют деятельность других нервных центров и рефлексов. Нейроны доминирующих центров приобретают более низкий уровень критической деполяризации мембран, т.е. становятся более возбудимыми, и способны эффективнее осуществлять пространственную и временную суммацию нервных импульсов. Синаптическое проведение к этим нейронам облегчено и поэтому они могут возбуждаться и за счет «посторонних» импульсов от не имеющих прямых связей с доминирующими центрами информационных каналов. Вследствие суммации многочисленных ВПСП (возбуждающий постсинаптический потенциал) возбуждение нейронов и число возбужденных клеток в доминирующем центре нарастает и осуществляемые им рефлекторные реакции легко реализуются. Преобладание рефлексов доминирующего центра над другими рефлекторными актами становится особенно выраженным - через систему вставочных нейронов доминирующий центр сопряженно тормозит другие центры и текущие рефлексы. Принцип доминанты позволяет концентрировать внимание и строить поведение для достижения определенной намеченной цели)
6)индукция (возникновение нервного процесса в определенном нервном центре под влиянием активности другого нервного центра. Выделяют следующие формы индукции: одновременную (положительную и отрицательную), последовательную (положительную и отрицательную). При одновременной положительной индукции происходит усиление активности взаимодействующих систем. Одновременная отрицательная индукция означает, что усиленная активность данного центра одновременно усиливает состояние торможения другого нервного центра. Доминантный очаг оказывает торможение других центров по механизму одновременной отрицательной индукции. Последовательная индукция проявляется в усилении активности ранее заторможенного центра (последовательная положительная индукция), либо в торможении ранее возбужденного центра (последовательная отрицательная индукция)).