Материал: Normalnaya_fiziologia_ekzamen2021
Виды мышечной ткани, их свойства и функции. Гетерогенность миоцитов скелетных мышц.
Виды мышечной ткани:
1)гладкая мышечная ткань. Состоит из одноядерных клеток — миоцитов веретеновидной формы длиной 20—500 мкм. Их цитоплазма в световом микроскопе выглядит однородно, без поперечной исчерченности. Обладает особыми свойствами: медленно сокращается и расслабляется, обладает автоматией, является непроизвольной (ее деятельность не управляется по воле человека). Входит в состав стенок внутренних органов: кровеносных и лимфатических сосудов, мочевыводящих путей, пищеварительного тракта (сокращение стенок желудка и кишечника);
2)поперечнополосатая скелетная мышечная ткань. Состоит из миоцитов, имеющих большую длину (до нескольких сантиметров) и диаметр 50—100 мкм; эти клетки многоядерные, содержат до 100 и более ядер; в световом микроскопе цитоплазма выглядит как чередование тёмных и светлых полосок. Свойства - высокая скорость сокращения, расслабления и произвольность (её деятельность управляется по воле человека). Входит в состав скелетных мышц, стенки глотки, верхней части пищевода, ей образован язык, глазодвигательные мышцы. Волокна длиной от 10 до 12 см;
3)поперечнополосатая сердечная мышечная ткань. Состоит из 1 или 2-х ядерных кардиомиоцитов, имеющих поперечную исчерченность цитоплазмы (по периферии цитолеммы). Кардиомиоциты разветвлены и образуют между собой соединения — вставочные диски, в которых объединяется их цитоплазма. Существует также другой межклеточный контакт - анастомозы (впячивание цитолеммы одной клетки в цитолемму другой). Этот вид мышечной ткани образует миокард сердца. Особое свойство - автоматия — способность ритмично сокращаться и расслабляться под действием возбуждения, возникающего в самих клетках (типичные кардиомиоциты). Эта ткань является непроизвольной (атипичные кардиомиоциты). Существует 3-й вид кардиомиоцитов - секреторные кардиомиоциты (в них нет фибрилл). Они синтезируют гормон тропонин, понижающий артериальное давление и расширяющий стенки кровеносных сосудов.
Мышечная оболочка сердца образована мышечными полостными клетками, которые с помощью вставочных дисков соеденяются в мышечные комплексы или сердечные мышечные волокна. Мышечная ткань имеет свойства восстанавливаться, она защищена соеденительной тканью, образующий рубец.
Мышечные волокна имеют поперечную исчерченность: темные анизотропные диски А (полоски А) чередуются со светлыми изотропным дисками I (полосками I). Диск А разделен светлой зоной (полосой Н), в центре которой проходит мезофрагма (М-линия). Диск И разделен темной Z-линией (телофрагма). Мышечные волокна содержат сократительные элементы - миофибриллы, состоящие из толстых (миозиновых) миофиламентов диаметром 10-15 нм и длиной 1,5 мкм, соответствующие дискам А, и тонких (актиновых) микрофиламентов диаметром 5-8 нм и длиной 1 мкм, соответствующие дискам I и прикрепляющиеся к телофрагме. Участок миофибриллы, расположенной между двумя телофрагмами, - саркомер (сократительная единица, длина которой примерно 2,5 мкм).
Свойства мышечной ткани:
1)возбудимость;
2)проводимость;
3)сократимость;
4)лабильность.
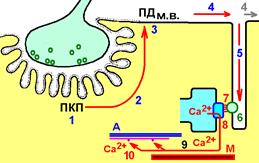
Электромеханическое сопряжение в миоците скелетной мышцы.
Совокупность явлений, обусловливающих связь между возбуждением (ПД) и сокращением мышечных волокон – электромеханическое сопряжение.
К регулярно расположенным Т‑трубочкам с двух сторон подходят терминальные цистерны саркоплазматического ретикулума (САР).
Т‑трубочка и расположенные с двух сторон от неё цистерны образуют триаду.
Z‑мембраны и зоны перекрытия актиновых и миозиновых нитей расположены рядом с триадами.
Такая структура необходима для эффективного электромеханического сопряжения.
Электромеханическое сопряжение включает следующие процессы:
1)образование потенциала концевой пластинки (ПКП);
2)электротоническое распространение ПКП в околосинаптическую область;
3)генерация потенциала действия мышечного волокна в околосинаптической области;
4)распространение потенциала действия по поверхностной мембране мышечного волокна;
5)распространение потенциала действия по мембранам поперечных трубочек (Т‑системы);
6)реакция рецепторов дигидропиридина на изменение мембранного потенциала;
7)передача сигнала дигидропиридиновыми рецепторами рианодиновым рецепторам терминальных цистерн саркоплазматического ретикулума;
8)открытие каналов Ca2+ с рианодиновыми рецепторами и выход Ca2+ из саркоплазматического ретикулума (цистерн) в саркоплазму (цитозоль);
9)диффузия Ca2+ в межфибриллярное пространство;
10)связывание Ca2+ с тропонином, расположенным на тонком, актиновом, миофиламенте;
11)конформационные изменения тропонина при связывании Са2+ => «заталкивание» тропомиозина в канавки актиновых нитей.
Типы и режимы мышечного сокращения.
Режимы мышечного сокращения:
1)изометрический – длина мышцы постоянна, но развивается внутреннее напряжение за счет силы растяжения упругих элементов (например, при сопротивлении давлению);
2)изотонический – укорочение мышцы при постоянном напряжении или нагрузке (сохранятся то же напряжение, например, при поднятии груза);
3)ауксотонический (смешанный) – длина мышцы уменьшается наряду с возрастанием развиваемой ею силы (меняется и напряжение, и длина мышцы).
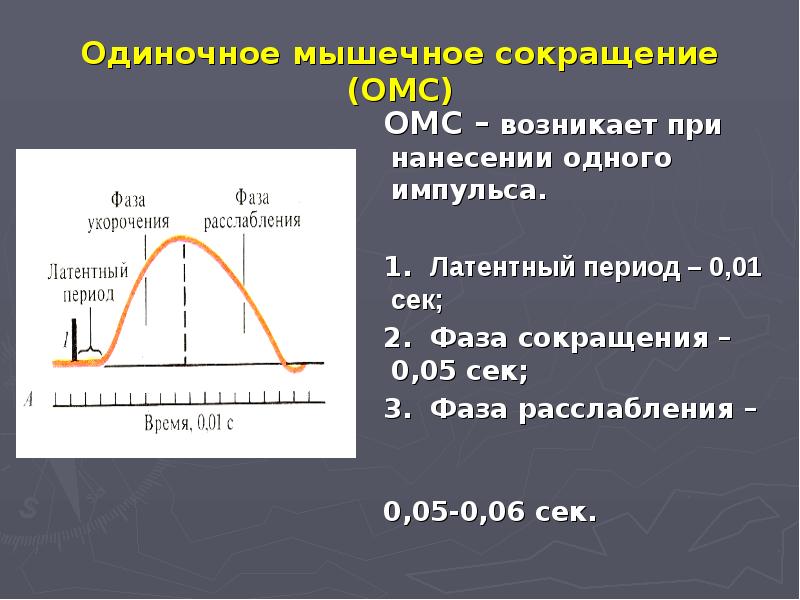
Существует два вида мышечных сокращений – одиночное и тетаническое. Одиночное мышечное сокращение является единственным видом сокращений для сердечной мышцы, а в скелетной мускулатуре оно носит искусственную этиологию и возникает в ответ на одиночный электрический сигнал и возникновение потенциала действия. Такое сокращение, длящееся 100 мс, имеет форму волны и включает три фазы:
1)латентный период (от 2-3 до 10 мс), длящийся от момента нанесения раздражения до начала сокращения;
2)фаза укорочения или сокращения (40-50 мс);
3)фаза расслабления (около 50мс).
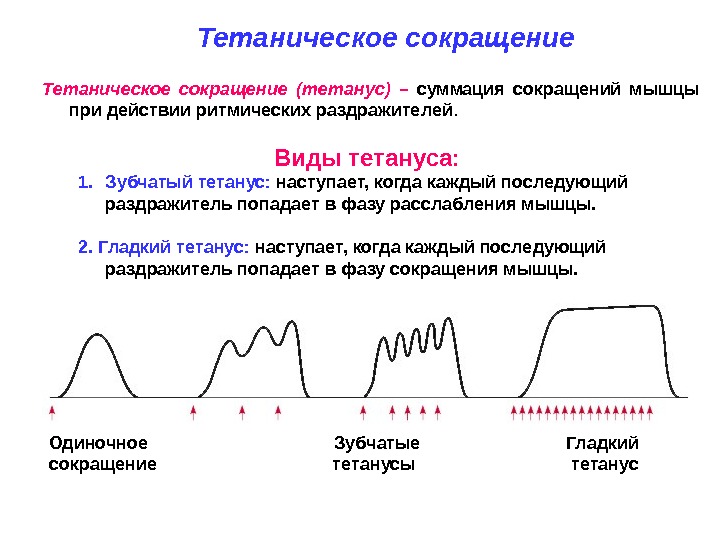
В естественных условиях импульсы поступают не одиночно, а сериями не менее 15-50 имп/с, на что мышца отвечает возникновением тетанического сокращения (тетануса). В его основе лежит явление суммации нескольких одиночных сокращений. В зависимости от частоты импульсов различают зубчатый и гладкий тетанус.
Зубчатый тетанус (неполный) возникает в том случае, когда каждый последующий импульс приходит в фазу расслабления мышцы.
Если частота раздражения выше, и каждый последующий импульс приходит в фазу укорочения мышцы, то происходит полная суммация, и тетаническое сокращение носит слитный характер – гладкий тетанус (полный).
Увеличение ответа при действии субмаксимальных раздражителей до определенного (максимального) уровня происходит за счет вовлечения в процесс возбуждения новых, не задействованных ранее, волокон. В случае дальнейшего возрастания раздражения (сверхмаксимальный уровень), ответ уже не увеличивается, и наоборот, при очень сильных раздражителях (5-10 и более порогов), можно достичь пессимального ответа.
В целостном организме мотонейроны посылают пачки потенциалов действия к двигательным единицам, которые в ответ сокращаются тетанически. Скелетные мышцы находятся в состоянии постоянного тонуса вследствие постоянной фоновой импульсации из моторных зон ЦНС.
Работу мышц делят на динамическую и статическую. Динамическая выполняется при перемещении груза. При динамической работе изменяется длина мышцы и ее напряжение. Следовательно мышца работает в ауксотоническом режиме. При статической работе перемещения груза не происходит, т.е. мышца работает в изометрическом режиме.
Классификация нервных волокон. Механизмы и законы проведения возбуждения по нервным волокнам.
Нервные волокна - отростки нейронов, которые покрыты глиальными оболочками. Оболочки нервных волокон в различных отделах нервной системы существенно разнятся по своему строению, что делит все волокна на безмиелиновые и миелиновые. Но все они состоят из отростка нервной клетки, находящегося в центре волокна, и имеют название осевого цилиндра (аксон), а также из глиальной оболочки, окружающей его.
1)нервные волокна типа А делятся на четыре подгруппы: α, β, γ и δ. Они покрыты миелиновой оболочкой. Наиболее толстые из них — так называемые альфа-волокна (Аα) — у теплокровных животных и человека имеют диаметр 12—22 мк и характеризуются значительной скоростью проведения возбуждения: 70—120 м/с. Такие волокна проводят возбуждение от моторных нервных центров спинного мозга к скелетным мышцам («двигательные» волокна) и от рецепторов мышц к соответствующим нервным центрам. Потенциал действия волокон Аα у теплокровных длится 0,5 мс. После его окончания развивается следовой отрицательный потенциал, который продолжается 15—20 мс и переходит в следовой положительный потенциал длительностью около 40—60 мс.
- Три другие группы волокон типа А: бета (Aβ)-, гамма (Аγ)- и дельта волокна имеют меньший диаметр — от 2 до 12 мк, меньшую скорость проведения и более длительный потенциал действия. Это преимущественно чувствительные волокна, проводящие возбуждение от различных рецепторов (тактильных, некоторых болевых, температурных и рецепторов внутренних органов) в ЦНС. Исключение составляют гамма-волокна, значительная часть которых проводит возбуждение в центробежном направлении — от клеток спинного мозга к интрафузальным мышечным волокнам, в которых расположены чувствительные нервные окончания — мышечные веретена.
2)к волокнам типа В относятся миелиновые, преимущественно преганглионарные волокна автономной нервной системы. Скорость проведения возбуждения в этих волокнах у теплокровных животных составляет 3—14 м/с. Продолжительность потенциала действия волокон типа В примерно в 2 раза превышает длительность потенциала действия волокон типа А. В среднем она составляет 1,2 мс. Отличительная особенность - в них не обнаруживается следового отрицательного потенциала: фаза реполяризации непосредственно переходит в следовой положительный потенциал, имеющий большую длительность — 100—300 мс.
3)к волокнам типа С относят безмякотные нервные волокна очень малого диаметра (порядка 0,5 мк). Скорость проведения возбуждения в этих волокнах 2—6 м/с. Большинство С-волокон относится к постганглионарным волокнам симпатической нервной системы. К С-волокнам относят также нервные волокна, которые участвуют в проведении возбуждения от болевых рецепторов и некоторых рецепторов холода, тепла и давления. Потенциалы действия С-волокон характеризуются наибольшей продолжительностью (2 мс у теплокровных животных). Имеют длительный следовой отрицательный потенциал (50—80 мс), сопровождающийся еще более продолжительной следовой позитивностью (300—1000 мс).
Проведение возбуждения по нервам подчиняется следующим законам:
1)закон анатомической и физиологической целостности нерва. Первая нарушается при перерезке, вторая – действии веществ, блокирующих проведение, например новокаина.
2)закон двустороннего проведения возбуждения. Распространяется в обе стороны от места раздражения. В организме чаще всего возбуждение по афферентным путям идет к нейрону, а по эфферентным – от нейрона. Такое распространение называется ортодромным. Очень редко возникает обратное (антидромное) распространение возбуждения.
3)закон изолированного проведения. Возбуждение не передается с одного нервного волокна на другое, входящее в состав этого же нервного ствола.
4)закон бездекрементного проведения. Возбуждение проводится по нервам без декремента, т.е. затухания. Следовательно, нервные импульсы не ослабляются, проходя по ним.
5)скорость проведения прямо пропорциональна диаметру нерва.
Механизм проведения возбуждения по нервным волокнам зависит от их типа. Существуют два типа нервных волокон: миелиновые и безмиелиновые.
Процессы метаболизма в безмиелиновых волокнах не обеспечивают быструю компенсацию расхода энергии. Распространение возбуждения будет идти с постепенным затуханием (декрементом). Декрементное поведение возбуждения характерно для низкоорганизованной нервной системы. Возбуждение распространяется за счет малых круговых токов, которые возникают внутрь волокна или в окружающую его жидкость. Между возбужденными и невозбужденными участками возникает разность потенциалов, которая способствует возникновению круговых токов. Ток будет распространяться от «+» заряда к «—». В месте выхода кругового тока повышается проницаемость плазматической мембраны для ионов Na, в результате чего происходит деполяризация мембраны. Между вновь возбужденным участком и соседним невозбужденным вновь возникает разность потенциалов, что приводит к возникновению круговых токов. Возбуждение постепенно охватывает соседние участки осевого цилиндра и так распространяется до конца аксона.
В миелиновых волокнах благодаря совершенству метаболизма возбуждение проходит, не затухая, без декремента. За счет большого радиуса нервного волокна, обусловленного миелиновой оболочкой, электрический ток может входить и выходить из волокна только в области перехвата. При нанесении раздражения возникает деполяризация в области перехвата А, соседний перехват В в это время поляризован. Между перехватами возникает разность потенциалов, и появляются круговые токи. За счет круговых токов возбуждаются другие перехваты, при этом возбуждение распространяется сальтаторно, скачкообразно от одного перехвата к другому. Сальтаторный способ распространения возбуждения экономичен, и скорость распространения возбуждения гораздо выше (70—120 м/с), чем по безмиелиновым нервным волокнам (0,5–2 м/с).