Материал: Normalnaya_fiziologia_ekzamen2021
Кривая "сила-длительность". Реобаза. Хронаксия. Лабильность.
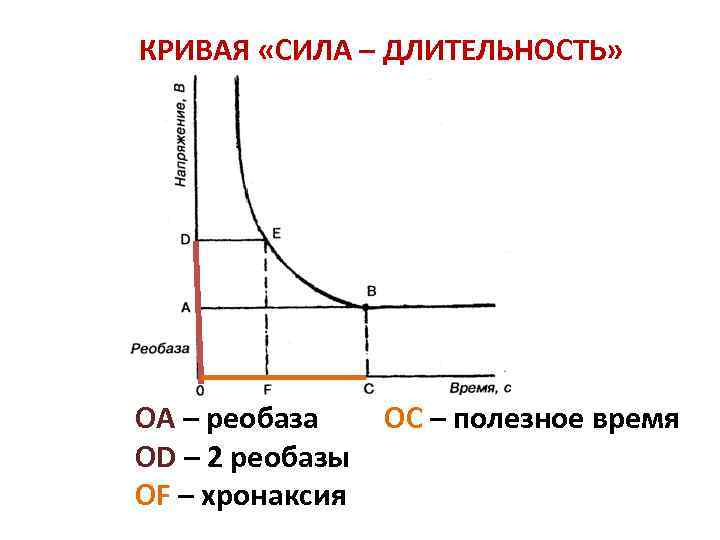
Кривая «сила-длительность» показывает соотношение между временем действия раздражителя и его амплитудой. На кривой видно, что уменьшение значения тока ниже определенной критической величины не приводит к возбуждению ткани независимо от продолжительности времени, в течение которого действует этот раздражитель.
Реобаза – минимальная величина тока, способная вызвать возбуждение при неограниченно длительном его действии. Реобаза - наименьшая сила постоянного электрического тока, вызывающая при достаточной длительности его действия возбуждение в живых тканях. Реобаза даёт представление о возбудимости тканей и органов по порогу силы и длительности действия раздражения. Реобаза соответствует порогу раздражения и выражается в вольтах или миллиамперах. Значение реобазы можно вычислить по формуле: i = a/t + b, где i - сила тока, t - длительность его действия, а и b - константы, определяемые свойствами ткани. Константа b является Р., так как при длительном действии раздражающего тока отношение a/t будет очень мало и i практически равняется b. Величина реобазы определяется разностью между критическим потенциалом и мембранным потенциалом покоя.
Хронаксия – минимальное время, в течение которого должен действовать ток, равный двум реобазам, чтобы вызвать ответную реакцию. Хронаксия - наименьшее время действия на ткань постоянного электрического тока удвоенной пороговой силы (удвоенной реобазы), вызывающего возбуждение ткани. Было также экспериментально установлено, что величина стимула, вызывающего возбуждающий эффект в тканях, находится в обратной зависимости от длительности его действия и графически выражается гиперболой - кривая <сила - время. Чем короче хронаксия, тем выше возбудимость.
Лабильность (Н. Е. Введенский) - свойство возбудимой ткани воспроизводить максимальное число потенциалов действия в единицу времени. Максимальная лабильность - у нервной ткани. Частота раздражений, вызывающая максимальную реакцию, - оптимальная, а вызывающая угнетение реакции - пессимальная. Мера лабильности - количество ПД, которое способно генерировать в единицу времени.
Реакции невозбудимых и возбудимых мембран на раздражители, градуальность и закон "всё или ничего".
Под действием многих раздражителей изменяется уровень мембранного потенциала. Такая реакция невозбудимых мембран связана, как правило, с изменением их ионной проницаемости за счет открывания или закрывания неспецифических потенциалнезависимых каналов. Между сдвигом мембранного потенциала невозбудимой мембраны и интенсивностью раздражителя, вызвавшего его, существует пропорциональная зависимость. Это свойство получило название градуальности. Оно присуще невозбудимой мембране во всем диапазоне изменений разности потенциалов на ней. Градуальность свойственна и возбудимой мембране, но только в том случае, если трансмембранная разность потенциалов, изменяясь под действием раздражителя, не достигает критического мембранного потенциала.
Такие раздражители, не доводящие деполяризацию до критического мембранного потенциала, и, следовательно, не вызывающие потенциала действия, называются подпороговыми для возбудимой ткани. Стимул, вызывающий сдвиг мембранного потенциала до КМП, считается пороговым, поскольку под действием его возникает потенциал действия (возбуждение).
Раздражитель может иметь разную природу (механическую, химическую, электрическую и т. д.), но пороговым он будет тогда, когда сдвинет уровень мембранного потенциала от потенциала покоя до критического мембранного потенциала: Un = |ПП| - |КМП|. Надпороговые (более сильные) раздражители возбудимой мембраны тем более вызывают потенциал действия.
Градуальность характерна и для отклонений трансмембранной разности потенциалов (от уровня потенциала покоя) в сторону, противоположную деполяризации (в аксоне кальмара от -85 до -90 мВ и более). Такой сдвиг мембранного потенциала называют гиперполяризацией. Градуальность присуща возбудимой мембране при любой ее гиперполяризации и при подпороговой (до критического мембранного потенциала) деполяризации.
Иллюстрацией сказанному служит реакция возбудимой мембраны на пропускание через нее слабого постоянного электрического тока. Под катодом развивается деполяризация, а под анодом — гиперполяризация. Гиперполяризационные сдвиги мембранного потенциала равны по абсолютной величине. Они тем больше, чем сильнее пропускаемый ток. Однако градуальность деполяризационных ответов под катодом свойственна только подпороговым электрическим стимулам, не приводящим к возбуждению. Не вызывая возбуждения, подпороговый электрический ток изменяет возбудимость возбудимой мембраны. Изменения возбудимости под действием подпорогового электрического тока - электротонические явления.
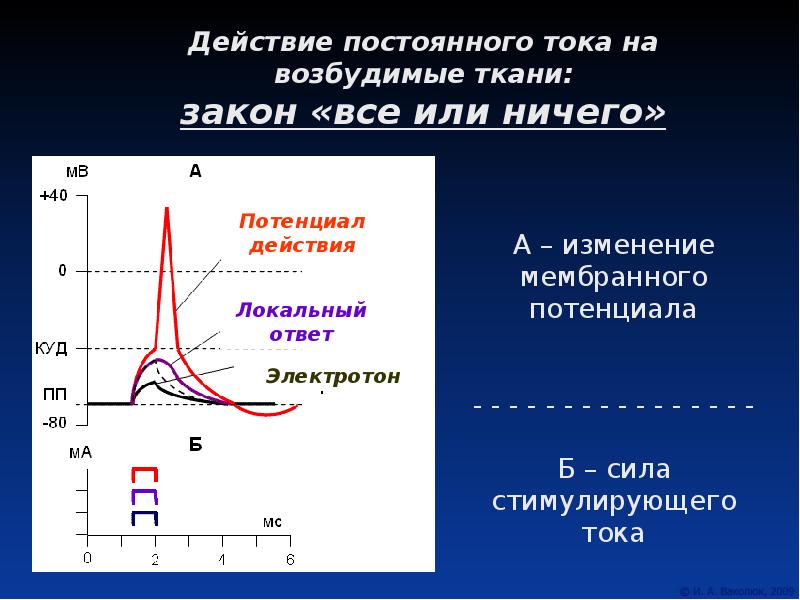
Под действием порогового и надпорогового раздражителей возбудимая мембрана генерирует ПД. Для этого процесса характерен закон «все или ничего». Смысл закона в том, что параметры потенциала действия (амплитуда, длительность, фронты) не зависят от интенсивности раздражителя. Как только достигается критический мембранный потенциал, изменения разности потенциалов на возбудимой мембране определяются только свойствами ее потенциалзависимых ионных каналов, которые обеспечивают входящий ток (из межклеточной среды в цитоплазму). Среди них внешний стимул открывает только самые чувствительные. Другие открываются за счет предыдущих, уже независимо от раздражителя. Говорят о спонтанном (самопроизвольном) характере процесса вовлечения в трансмембранный перенос ионов все новых потенциалзависимых ионных каналов. Поэтому амплитуда, длительность, крутизна переднего и заднего фронтов потенциала действия зависят только от ионных градиентов на клеточной мембране и кинетических характеристик ее каналов.
Закон «все или ничего» — характернейшее свойство одиночных клеток и волокон, обладающих возбудимой мембраной. Большинству многоклеточных образований он не свойствен, поскольку разные клетки даже в одном органе имеют неодинаковый порог возбуждения. Чем больше клеток вовлекают в возбуждение, тем сильнее ответ многоклеточной структуры. Исключение составляют структуры, организованные по типу синцития. Их возбуждение подчиняется закону «все или ничего».
Закон «все или ничего»: подпороговый раздражитель вызывает местную деполяризацию («ничего»), пороговый раздражитель вызывает максимально возможный ответ («все»). Сверхпороговый раздражитель вызывает такой же ответ, что и пороговый.
Первичные и вторичные электротонические явления. Использование их в медицине.
Электротонические явления развиваются в нервной и мышечной тканях при пропускании через них постоянного подпорогового электрического тока. Под катодом возникает деполяризация, а под анодом - гиперполяризация. Де- и гиперполяризационные сдвиги мембранного потенциала равны по абсолютной величине. Они тем больше, чем сильнее пропускаемый ток. Не вызывая возбуждения, подпороговый электрический ток изменяет возбудимость возбудимой мембраны. Изменения возбудимости под действием подпорогового электрического тока называют электротоническим явлением (точнее, первичным электротоническим явлением). Обнаружил Э. Пфлюгер в 1859 г.
Количественной мерой возбудимости служит интенсивность порогового раздражителя. Её можно выразить значением напряжения (Uпор) U=|ПП|-|КМП|. Под катодом уровень мембранного потенциала приближен к КМП. На этом фоне переход возбудимой мембраны в возбужденное состояние происходит под действием более слабого порогового раздражителя => возбудимость под катодом повышается. Под анодом уровень мембранного потенциала отдален от КМП Uпа>Uп0, => возбудимость под анодом уменьшается. Повышение возбудимости под катодом называется катэлектротоном, а понижение её под анодом – анэлектротоном. Анэлектротон и катэлектротон в совокупности составляют первичные электротонические явления. В них проявляется возможность управления возбудимостью тканей организма при помощи подпорогового электрического тока, который изменяет уровень ПП, но не сдвигает КМП. Это физиотерапевтическая процедура называется гальванизация.
При продолжительном действии тока живая ткань реагирует на него извращением возбудимости: катэлектротон сменяется катодической депрессией (понижением возбудимости под катодом), а анэлектротон - анодической экзальтацией (повышением возбудимости под анодом). При неправильном проведении процедуры гальванизации можно достичь под анодом не ослабления боли, а усиления её. Это так называемые вторичные электротонические явления (Б.Ф. Вериго, 1883). Механизмом катодической депрессии является ослабление проницаемости потенциалзависимых натриевых каналов и усиление проницаемости калиевых каналов. Вторичные электротонические явления обусловлены действием постоянного подпорогового электрического тока на КМП.

Рефрактерность (фазы). Количественная мера возбудимости.
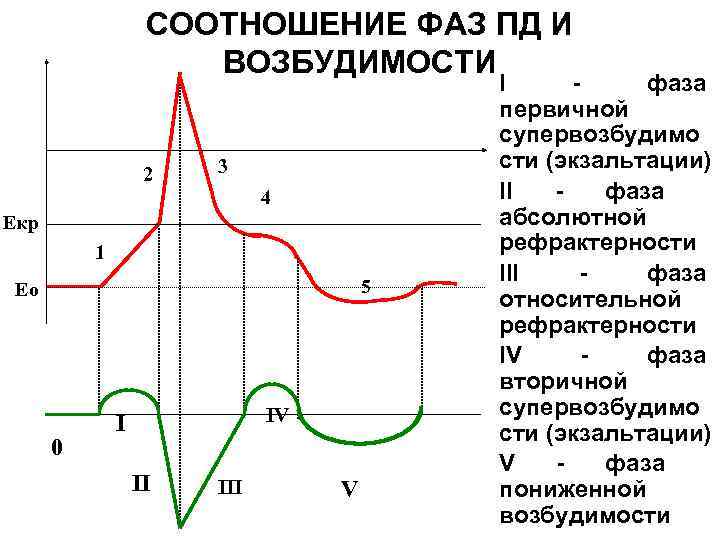
Рефрактерность - временное снижение возбудимости ткани, возникающее при появлении потенциала действия. В этот момент повторные раздражения не вызывают ответной реакции (абсолютная рефрактерность). Она длится не более 0,4 мс, затем наступает фаза относительной рефрактерности, когда раздражение может вызвать слабую реакцию. Эта фаза сменяется фазой повышенной возбудимости - супернормальности. Показатель рефрактерности (рефрактерный период) - время, в течение которого возбудимость ткани снижена. Рефрактерный период тем короче, чем выше возбудимость ткани.
Процесс возбуждения сопровождается изменением возбудимости. Таков смысл свойства рефрактерности (ввел в науку Э.Ж. Марей, в 1876 г. - угнетение возбудимости миокарда в момент его возбуждения. 1908 г., Н.Е. Введенский - вслед за угнетением наступает некоторое повышение возбудимости возбужденной ткани).
Выделяют три основные стадии рефрактерности, их принято называть фазами:
1)развитие возбуждения вначале сопровождается полной утратой возбудимости (е = 0). Это состояние - абсолютно рефрактерная фаза. Она соответствует времени деполяризации возбудимой мембраны. В течение абсолютно рефрактерной фазы возбудимая мембрана не может генерировать новый потенциал действия, даже если на нее действовать сколь угодно сильным раздражителем (S„-> оо). Природа абсолютно рефрактерной фазы состоит в том, что во время деполяризации все потенциалзависимые ионные каналы находятся в открытом состоянии, и дополнительные стимулы не могут вызвать воротный процесс (им просто не на что действовать).
2)относительно рефрактерная фаза – возвращает возбудимость от нуля к исходному уровню (е0). Относительно рефрактерная фаза совпадает с реполяризацией возбудимой мембраны. С течением времени во все большем числе потенциалзависимых ионных каналов завершаются воротные процессы, с которыми было связано предшествующее возбуждение, и каналы вновь обретают способность к следующему переходу из закрытого в открытое состояние под действием очередного стимула. Во время относительно рефрактерной фазы пороги возбуждения постепенно снижаются (S„o<S„<со) и, следовательно, возбудимость восстанавливается до исходного уровня (0<е<е0).
3)фаза экзальтации, для которой характерна повышенная возбудимость (е>е0). Она связана с изменением свойств сенсора напряжения во время возбуждения. За счет перестройки конформации белковых молекул изменяются их дипольные моменты, что приводит к повышению чувствительности сенсора напряжения к сдвигам мембранного потенциала (критический мембранный потенциал приближается к потенциалу покоя).
Разным возбудимым мембранам присуща неодинаковая продолжительность каждой фазы рефрактерности. Так, в скелетных мышцах АРФ длится в среднем 2,5 мс, ОРФ — около 12 мс, ФЭ — приблизительно 2 мс. Миокард отличается гораздо более продолжительной АРФ — 250—300 мс, что обеспечивает четкую ритмичность сердечных сокращений и является необходимым условием жизни. В типичных кардиомиоцитах относительно рефрактерная фаза длится около 50 мс, а в сумме продолжительность абсолютно рефрактерной и относительно рефрактерной фаз примерно равна длительности потенциала действия. Различия в длительности рефракторных фаз обусловлены неодинаковой инерционностью потенциалзависимых ионных каналов. В тех мембранах, где возбуждение обеспечивается натриевыми каналами, рефрактерные фазы наиболее быстротечны и потенциал действия наименее продолжителен (порядка единиц мс). Если за возбуждение ответственны кальциевые каналы (например, в гладких мышцах), то рефрактерные фазы затягиваются до секунд. В сарколемме кардиомиоцитов присутствуют и те, и другие каналы, вследствие чего длительность рефрактерных фаз занимает промежуточное значение (сотни мс).
В фазу деполяризации потенциала действия потенциалзависимые натриевые ионные каналы кратковременно открываются, но затем инактивируются h-ворота. В период инактивации натриевых ионных каналов возбудимые клетки не способны реагировать повышением натриевой проницаемости на повторный стимул. Поэтому во время фазы деполяризации мембрана не может генерировать потенциал действия в ответ на действие пороговых или сверхпороговых раздражителей. Это состояние называется абсолютной рефрактерностью, время которой составляет в нервных волокнах 0,5—1,0 мс, а в скелетных мышечных клетках —в среднем 2 мс. Период абсолютной рефрактерности заканчивается после того, как снижается количество инактивированных натриевых каналов и постепенно увеличивается количество натриевых каналов, находящихся в закрытом состоянии. Происходят эти процессы во время фазы реполяризации, когда уменьшению количества потенциалзависимых натриевых ионных каналов, находящихся в состоянии инактивации, соответствует период относительной рефрактерности. Период относительной рефрактерности характеризуется тем, что лишь некоторая часть потенциалзависимых натриевых ионных каналов переходит в закрытое состояние, а в силу этого порог возбудимости мембраны клетки имеет более высокие значения, чем в исходном состоянии. Поэтому возбудимые клетки в период относительной рефрактерности могут генерировать потенциалы действия, но при воздействии на них раздражителей сверхпороговой силы. Однако из-за небольшого количества потенциалзависимых натриевых ионных каналов, находящихся в закрытом состоянии, амплитуда генерируемых при этом потенциалов действия будет меньше, чем в условиях исходной возбудимости нервной или мышечной клетки.
В клетках возбудимых тканей максимальное количество генерируемых потенциалов действия в единицу времени обусловлено двумя факторами: длительностью потенциала действия и длительностью периода абсолютной рефрактерности после каждого импульса. На этом основании в физиологии формулируется понятие лабильности: чем меньше период абсолютной рефрактерности при возбуждении возбудимой ткани, тем выше ее функциональная подвижность или лабильность, тем больше в ней генерируется потенциалов действия в единицу времени.
При непрерывной стимуляции нерва электрическим током лабильность нерва зависит от частоты и силы раздражения. В зависимости от частоты и силы раздражения нерва сокращение иннервируемой им мышцы может быть максимальной или минимальной амплитуды. Эти явления были названы соответственно оптимумом и пессимумом (Н. Е. Введенский). Максимальное (оптимально большое) сокращение мышцы возникает в том случае, если каждый последующий электрический стимул действует на нерв в периоде его состояния супернормальной возбудимости после предыдущего потенциала действия. Минимальное (или пессимальное) сокращение мышцы возникает в том случае, если каждый последующий электрический стимул действует на нерв, находящийся в периоде относительной рефрактерности после предыдущего потенциала действия. Поэтому значения оптимальной частоты раздражения нерва всегда меньше, чем значения пессимальной частоты раздражения.
К мерам возбудимости относятся:
1)порог раздражения - первая базисная мера раздражителя любой природы, но для количественной оценки возбудимости в медицине используют не любой раздражитель, а электрический ток. Именно с помощью электрического тока тестируют мышцы, нервы, синапсы. Электрический ток точно дозируется, причем по двум показателям: по силе и по времени действия. С другими раздражителями иначе: например, химический можно дозировать по силе (концентрации), но нельзя по длительности, т.к. для его отмывания нужно время. С помощью электрического тока получены еще 3 меры возбудимости, одна из которых используется в медицине:
2)базисная мера - реобаза - минимальная сила постоянного тока, которая, действуя длительное, но определенное время, способна вызвать ответную реакцию. Недостаток этой меры - определение времени сложно – расплывчато;
3)полезное время - то время, которое должна действовать сила тока в 1 реобазу, чтобы вызвать ответную реакцию. Но и эта мера возбудимости не нашла своего применения в медицинской практике, потому что она находится на очень пологой части кривой "сила - время" и любая неточность вела к большой ошибке;
4)хронаксия - минимальное время, в течение которого должна действовать сила тока в 2 реобазы, чтобы вызвать ответную реакцию. На графике - тот участок кривой, где зависимость между силой и временем точно прослеживается. Посредством хронаксии определяют возбудимость нервов, мышц, синапсов. Этим методом определяют, где наступило поражение нервно-мышечной системы: на уровне мышцы, нервов, синапсов или центральных образований.