Материал: Normalnaya_fiziologia_ekzamen2021
У человека в центральной части сетчатки расположены цветочувствительные рецепторы — нервные клетки, которые называются колбочками. Каждый из трёх видов колбочек имеет свой тип цветочувствительного пигмента белкового происхождения. Один тип пигмента чувствителен к красному цвету с максимальной длиной волны 552—557 нм, другой — к зелёному (максимум около 530 нм), третий — к синему (426 нм). Люди с нормальным цветным зрением имеют в колбочках все три пигмента (красный, зелёный и синий) в необходимом количестве. Их называют трихроматами (от др.-греч. χρῶμα — цвет).
Термин «цветовая слепота» не совсем четко отражает проблемы расстройства цветового зрения - люди, обладающие этой аномалией, видят все не в черно-белом цвете: они не различают некоторые цвета или их оттенки. Поэтому наиболее приемлемым назвать это «нарушением цветового зрения». В связи с этим существует несколько видов этого нарушения.
Наиболее часто встречающимся видом нарушения цветового зрения является «дейтераномалия», расстройство восприятия зеленого цвета. При дейтераномалии зелёный цвет смешивается со светло-оранжевым, светло-розовым. Люди с дейтераномальным зрением могут даже не знать о своей аномалии. При еще одной разновидности нарушения цветового зрения, называемой «протаномалией» (слабость восприятия красного цвета) красный цвет смешивается со светло-зелёным, светло-коричневым. Цветовая слепота в сине-фиолетовой области спектра называется «тританомалия»; она встречается крайне редко и практического значения не имеет. При тританомалии все цвета спектра представляются оттенками красного или зелёного.
Люди, различающие только два цвета из трех основных, обладают двухцветным зрением, что значительно более серьезнее, чем аномалия трихромазии, о которой рассказывалось выше. Двухцветное зрение бывает трех видов:
1)дейтеранопия – слепота на зеленый цвет (длинные волны);
2)протанопия – слепота на красный цвет (средние волны);
3)тританопия – цветовая слепота на синий цвет (короткие волны).
Монохромазия – еще одна разновидность нарушения цветового восприятия. Монохроматы видят все в черном и белом цветах и оттенках серого. Различают два вида монохромазии: монохромазия палочки (клетка сетчатки глаза) и монохромазия колбочки сетчатки. Первый вид цветовой слепоты также называют ахроматопсией. При этом виде нарушения люди страдают плохим зрением и высокой чувствительностью к свету. У некоторых может развиться нистагм (непроизвольные ритмические двухфазные движения глазных яблок).
Рецептивные поля сетчатки. Острота зрения. Поле зрения.
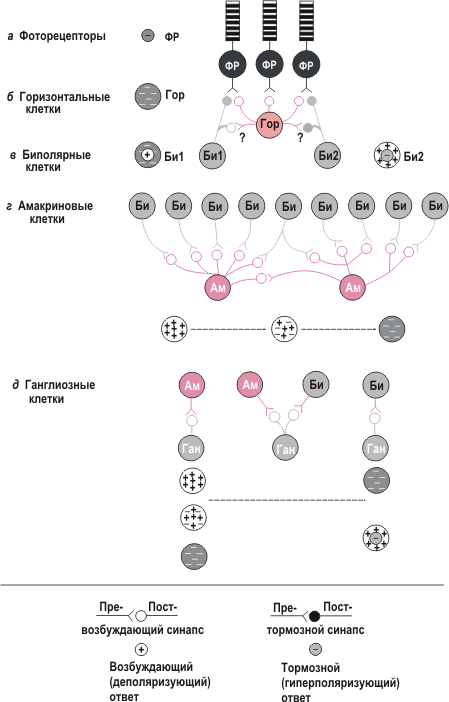
Активация рецептивных полей ганглиозных клеток сетчатки - важный этап переработки зрительной информации, поскольку от него зависят характеристики зрительных сигналов, направляемых в мозг.
Рецептивное поле фоторецептора обозначено на рисунке а маленьким кружком со знаком минус. Маленький кружок соответствует его небольшим размерам и округлым очертаниям рецептивного поля, а знак минус - гиперполяризационной реакции фоторецепторной клетки на его освещение.
Рецептивное поле горизонтальной клетки на рисунке б представлено в виде кружка большего размера со знаком минус. При попадании света на один из фоторецепторов, конвергирующих (сходящихся) на данной клетке, та гиперполяризуется. Последовательность событий:
1)свет вызывает гиперполяризацию одного или нескольких фоторецепторов;
2)уменьшается высвобождение нейромедиатора из окончаний этих фоторецепторов;
3)горизонтальная клетка гиперполяризуется вследствие ослабления тонического возбуждающего влияния.
Рецептивные поля биполярных клеток двух типов показаны на рисунке в. Рецептивное поле биполярной клетки слева (Би1) состоит из центральной возбуждающей области (белый кружок со знаком плюс), которая окружена тормозной рецептивной областью (серое кольцо со знаком минус). Организованное подобным образом, оно называется центрально-периферическим; в данном случае это - рецептивное поле с on-центром и off-периферией (для краткости "с on-центром"). Биполярная клетка справа (Би2) имеет рецептивное поле с off-центром и on-периферией (для краткости "с off-центром"): центральная тормозная область ее рецептивного поля (знак минус) окружена возбуждающей периферической (знак плюс).
Ответы биполярных клеток определяются тем, каков их вход: либо короткий - от одного или нескольких фоторецепторов, либо более длинный - через горизонтальные клетки. Ответ на стимуляцию центра рецептивного поля осуществляется в результате прямых связей клетки с одним или несколькими фоторецепторами. Если нейромедиатор, тонически высвобождаемый из синаптических окончаний фоторецептора, гиперполяризует биполярную клетку, то при гиперполяризации фоторецептора световым стимулом количество высвобождаемого нейромедиатора уменьшится и произойдет ее деполяризация (растормаживание). С другой стороны, если тонически высвобождаемый нейромедиатор является деполяризующим, световой стимул приводит к гиперполяризации биполярной клетки (снятию облегчения, как в горизонтальной клетке на рисунке б). Световой стимул, попадающий на фоторецепторы периферии рецептивного поля, изменяет активность горизонтальных клеток - вход к биполярным клеткам обеспечивается через более длинный путь. Сигналы от горизонтальных клеток сопровождаются ответами биполярных, противоположными по знаку по сравнению с прямыми ответами на стимуляцию фоторецепторов центра рецептивного ноля.
В путях, связывающих фоторецепторы сетчатки с биполярными и горизонтальными клетками, нейромедиатором служит возбуждающая аминокислота (глутамат). Она деполяризует биполярные клетки с off-центром, а также горизонтальные в результате активации ионотропных глутаматных рецепторов постсинаптической мембраны. Биполярные клетки с on-центром та же кислота гиперполяризует, воздействуя на метаботропные глутаматные рецепторы.
При одновременном освещении фоторецепторов периферии и центра рецептивного поля биполярные клетки могут вообще не ответить из-за антагонистического характера влияний центра и периферии. При движении луча света через рецептивное поле их активность будет последовательно меняться по мере того, как луч проходит от периферии к центру и снова к периферии.
Рецептивные поля амакриновых клеток изображены на рисунке г. Сигналы к этим клеткам поступают при различных сочетаниях активности биполяров с on- и оff-центрами. Их рецептивные поля - разные сочетания областей с on- и оff-центрами. Существует много типов амакриновых клеток; известны, по крайней мере, восемь разных нейромедиаторов, высвобождаемых ими.
Рецептивные поля ганглиозных клеток показаны на рисунке д. Эти клетки получают доминирующий вход от амакриновых (левая клетка Ган), смешанный вход от амакриновых и биполярных (средняя клетка Ган) либо доминирующий вход от биполярных клеток (правая клетка Ган). Если доминирует вход от амакриновых клеток, рецептивное поле ганглиозных носит рассеянный характер - местами оно возбуждающее, местами тормозное. Иная ситуация при доминировании входа от биполярных клеток: рецептивное поле ганглиозных клеток организовано по принципу центр/периферия, как у биполярных.
Острота зрения - функция зрительной системы, позволяющая наблюдателю различать мелкие объекты внешнего мира. Характеризуется наименьшим углом зрения, под которым две рассматриваемые точки воспринимаются как раздельные. Количественно выражается величиной, обратной этому минимальному углу, выраженному в минутах. Зависит от освещенности, контрастности, формы объекта и других факторов.
В странах СНГ определяется долями единицы: 1,0 — нормальное зрение, 0,9; 0,8, и т. д. до 0,1 — определяется количеством строк начиная с верхней по таблице Сивцева или Головина с расстояния 5 метров. Исследования проводят для каждого глаза отдельно: сначала определяют остроту зрения одного, затем другого глаза.
При исследовании остроты зрения с другого расстояния (меньше 0,1 — если человек с 5 метров не распознает знаки верхнего ряда), проверяемого приближают к таблице и через каждые 0,5 метра спрашивают, пока он не назовёт правильно знаки верхнего ряда. Величина рассчитывается по формуле:
V = d / D, где V — острота зрения; d — расстояние, с которого проводится исследование; D — расстояние, на котором нормальный глаз видит данный ряд.
Поле зрения - пространство, которое видит глаз человека при фиксации взора в одной точке и неподвижной голове. В зависимости от того, участвуют в зрении один или оба глаза, различают монокулярное и бинокулярное поле зрения. Каждый глаз человека имеет в норме поле ахроматического зрения: 55° вверх, 60° вниз, 90° наружу (суммарное поле зрения двумя глазами – 180°) и 60° – внутрь. Ахроматическое поле больше хроматического, т.к. обусловлено деятельностью палочек, число которых больше и которые расположены преимущественно на периферии сетчатки. Для различных цветов поле зрения также неодинаково: наименьший размер поля зрения – у зелёного цвета, наибольший – у синего. В нормальном поле зрения имеются физиологические скотомы (слепые участки): слепое пятно Мариотта, что соответствует проекции диска зрительного нерва, не содержащего фоторецепторы.
Структурно-функциональная схема слухового анализатора.
Наружное ухо - звукоулавливающий аппарат. Наружный слуховой проход проводит звуковые колебания к барабанной перепонке. Барабанная перепонка, отделяющая наружное ухо от барабанной полости, или среднего уха, представляет собой тонкую (0,1 мм) перегородку, имеющую форму направленной внутрь воронки. Перепонка колеблется при действии звуковых колебаний, пришедших к ней через наружный слуховой проход.
Звуковые колебания улавливаются ушными раковинами (у животных они могут поворачиваться к источнику звука) и передаются по наружному слуховому проходу к барабанной перепонке, которая отделяет наружное ухо от среднего. Улавливание звука и весь процесс слушания двумя ушами — бинауральный слух — имеет значение для определения направления звука. Звуковые колебания, идущие сбоку, доходят до ближайшего уха на несколько десятитысячных долей секунды (0.0006 с) раньше, чем до другого. Этой ничтожной разницы во времени прихода звука к обоим ушам достаточно, чтобы определить его направление.
Среднее ухо - звукопроводящий аппарат. Оно представляет собой воздушную полость, которая через слуховую (Евстахиеву) трубу соединяется с полостью носоглотки. Колебания от барабанной перепонки через среднее ухо передают соединенные друг с другом 3 слуховые косточки — молоточек, наковальня и стремечко, последнеe через перепонку овального окна передает эти колебания жидкости, находящейся во внутреннем ухе, — перилимфе.
Благодаря особенностям геометрии слуховых косточек стремечку передаются колебания барабанной перепонки уменьшенной амплитуды, но увеличенной силы. Кроме того, поверхность стремечка в 22 раза меньше барабанной перепонки, что во столько же раз усиливает его давление на мембрану овального окна. В результате этого даже слабые звуковые волны, действующие на барабанную перепонку, способны преодолеть сопротивление мембраны овального окна преддверия и привести к колебаниям жидкости в улитке.
При сильных звуках специальные мышцы уменьшают подвижность барабанной перепонки и слуховых косточек, адаптируя слуховой аппарат к таким изменениям раздражителя и предохраняя внутреннее ухо от разрушения.
Благодаря соединению через слуховую трубу воздушной полости среднего уха с полостью носоглотки возникает возможность выравнивания давления по обе стороны барабанной перепонки, что предотвращает ее разрыв при значительных изменениях давления во внешней среде — при погружениях под воду, подъемах на высоту, выстрелах и пр. - барофункция уха.
В среднем ухе расположены две мышцы: напрягающая барабанную перепонку и стременная. Первая, сокращаясь, усиливает натяжение барабанной перепонки и тем самым ограничивает амплитуду ее колебаний при сильных звуках, а вторая фиксирует стремечко и тем самым ограничивает его движения. Рефлекторное сокращение этих мышц наступает через 10 мс после начала сильного звука и зависит от его амплитуды. Этим внутреннее ухо автоматически предохраняется от перегрузок. При мгновенных сильных раздражениях (удары, взрывы и т. д.) этот защитный механизм не успевает сработать, что может привести к нарушениям слуха (например, у взрывников и артиллеристов).
В перепончатом лабиринте волокна преддверно-улиткового нерва оканчиваются в нейроэпителиальных волосковых клетках (рецепторах), находящихся в определенных местах. Пять рецепторов относятся к вестибулярному анализатору, из них три расположены в ампулах полукружных каналов и называются ампулярными гребешками, а два находятся в мешочках и носят название пятен.
Один рецептор является слуховым, он располагается на основной мембране улитки и называется кортиевым (спиральным) органом. Во внутреннем ухе расположены рецепторы слухового и статокинетического анализаторов. Рецепторный (звуковоспринимающий) аппарат слухового анализатора находится в улитке и представлен волосковыми клетками спирального (кортиева) органа. Улитка и заключенный в ней рецепторный аппарат слухового анализатора называются кохлеарным аппаратом. Звуковые колебания, возникающие в воздухе, передаются через наружный слуховой проход, барабанную перепонку и цепь слуховых косточек на вестибулярное окно лабиринта, вызывают волнообразные перемещения перилимфы, которые, распространяясь, передаются на спиральный орган. Рецепторный аппарат статокинетического анализатора, расположенный в полукружных каналах и мешочках преддверия, - вестибулярный аппарат.
Проводниковый отдел слухового анализатора представлен перефирическим биполярным нейроном, расположенным в спиральном ганглии улитки (первый нейрон). Волокна слухового или (кохлеарного) нерва, образованные аксонами нейронов спирального ганглия, заканчиваются на клетках ядер кохлеарного комплекса продолговатого мозга (второй нейрон). После частичного перекреста волокна идут в медиальное коленчатое тело таламуса, где происходит переключение (третий нейрон), отсюда возбуждение поступает в кору (четвертый) нейрон. В медиальных (внутренних) коленчатых телах, а также в нижних буграх четверохолмия располагаются центры рефлекторных двигательных реакций, возникающих при действии звука.
Корковый (центральный) отдел слухового анализатора находится в верхней части височной доли большого мозга (верхняя височная) извилина, поля 41 и 42 по Бродману. Важное значение для функции слухового анализатора имеют поперечные височные обеспечивающими регуляцию деятельности всех уровней извилины Гешля. При двустороннем разрушении указанных полей - полная глухота. Когда поражение ограничивается одним полушарием, может наступить небольшое и нередко лишь временное понижение слуха. Это объясняется тем, что проводящие пути слухового анализатора неполностью перекрещиваются. Оба внутренних коленчатых тела связаны между собой промежуточными нейронами, через которые импульсы могут переходить с правой стороны на левую и обратно. В результате корковые клетки каждого полушария получают импульсы с обоих кортиевых органов.
Слуховая сенсорная система дополняется механизмами обратной связи, обеспечивающими регуляцию деятельности всех уровней слухового анализатора с участием нисходящих путей. Такие пути начинаются от клеток слуховой коры, переключаясь последовательно в медиальных коленчатых телах таламуса, задних (нижних) буграх четверохолмия, в ядрах кохлеарного комплекса. Входя в состав слухового нерва, центробежные волокна достигают волосковых клеток кортиева органа и настраивают их на восприятие опрелеленных звуковых сигналов.