Материал: Лекции Хомича Цитология
мок (плацента, жовте тіло яєчників). До складу цієї системи належать також поодинокі ендокриноцити, які є в більшості органів.
Нейросекреторні ядра гіпоталамуса. Гіпоталамус утворює нижню стінку проміжного мозку. Це центральний нейроендокринний орган, який поєднує нервову і гуморальну регуляцію більшості нутрощів організму. У ньому міститься близько 32 пар ядер (скупчення нейронів), частина яких утворена нейросекреторними клітинами. Ядра розташовані в передній, середній і задній зонах гіпоталамуса.
У п е р е д н і й з о н і гіпоталамуса містяться дві пари ядер:
супраоптичні та паравентрикулярні. Нейросекреторні клітини обох ядер продукують гормон вазопресин. Він стимулює скорочення гладких м’язових клітин стінки кровоносних судин, зумовлюючи цим підвищення тиску крові. Цей гормон сприяє реабсорбції води у нирках, що призводить до зменшення сечовиділення (антидіуретичний гормон), бере участь у регуляції температури тіла, серцево-судинної діяльності і необхідний для розвитку головного мозку.
Нейросекреторні клітини паравентрикулярних ядер продукують ще й гормон окситоцин, який викликає скорочення гладких м’язових клітин м’язової оболонки стінки матки і міоепітеліоцитів молочної залози.
Гормони нейросекреторних клітин ядер передньої зони гіпоталамуса стікають по аксонах цих клітин у нейрогіпофіз, де і потрапляють у кров.
Нейросекреторні клітини ядер с е р е д н ь о ї з о н и г і п о т а л а м у с а (аркуатне, супрахіазматичне, вентромедіальне, дорсальне тощо) продукують дві групи гормонів: ліберини і статини (релізингфактори).
Ліберини стимулюють продукування і виведення гормонів передньої частини аденогіпофіза, а статини, навпаки, пригнічують ці процеси.
Нейроцити ядер з а д н ь о ї з о н и г і п о т а л а м у с а і час-
тково ядер перших двох зон своїми відростками беруть участь у формуванні нервових стовбурів автономної нервової системи, які прямують до нутрощів.
Гіпофіз – центральний ендокринний орган. Він продукує гормони, які впливають на діяльність периферичних ендокринних залоз (гіпофіззалежних) і багатьох неендокринних клітин різних тканин та органів. У гіпофізі виділяють аденогіпофіз і нейрогіпофіз (рис. 47).
А д е н о г і п о ф і з розвивається з епітелію даху первинної ротової порожнини. Він утворений дистальною (передньою), проміжною і туберальною частинами. Ендокриноцити усіх частин формують тра-
168
бекули, між якими є ніжні прошарки пухкої волокнистої сполучної тканини з кровоносними судинами.
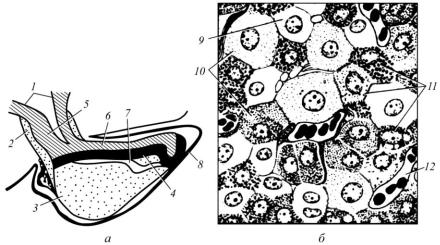
Рис. 47. Схема будови гіпофіза великої рогатої худоби (а) і ділянки дистальної частини аденогіпофіза (б):
1 – лійка; 2 – туберальна; 3 – дистальна; 4 – проміжна частини аденогіпофіза; 5 – стовбур лійки; 6 – нейрогіпофіз; 7 – щілина між дистальною і проміжною частинами аденогіпофіза; 8 – тверда мозкова оболонка; 9 – хромофобні, 10 – ацидофільні; 11 – базофільні аденоцити; 12 – кровоносний капіляр
Дистальна частина аденогіпофіза сформована хромофільними
(40 %) і хромофобними ендокриноцитами (60 %).
Хромофільні ендокриноцити містять у цитоплазмі гранули, які інтенсивно фарбуються гістологічними барвниками. Одні з них фарбуються основними барвниками – базофільні, інші – кислими – ацидофільні. Серед ацидофільних ендокриноцитів виділяють соматотропоцити і лактотропоцити. Соматотропоцити мають кулясті секреторні гранули діаметром 200–400 нм і продукують соматотропін, який стимулює ріст організму. Секреторні гранули лактотропоцитів овальні. Їх довжина – 500–600 нм, а ширина 100–200нм. Лактотропоцити продукують пролактин, який стимулює секрецію молока.
До базофільних ендокриноцитів належать гонадотропоцити, тиротропоцити і кортикотропоцити. Гонадотропоцити крупні, діаметр їх секреторних гранул становить 200–300 нм. В їх центральній частині є пляма, яка утворена розширеними порожнинами цистерн комплексу Гольджі. Гонадотропоцити продукують два гормони: фолітропін і лютропін. Перший стимулює сперматогенез і овогенез та секрецію естрогену, а другий впливає на розвиток жовтого тіла яєчників і стимулює секрецію тестостерону в самців. Тиротропоцити мають дрібні секреторні гранули (80–150 нм). Вони синтезують тиротропін, який стимулює синтез і секрецію гормонів щитоподібної залози. Кортикотропоцити мають секреторні гранули у вигляді пухирців із щільною
169
серцевиною. Ці клітини продукують адренокортикотропін, який стимулює секрецію гормонів кіркової речовини надниркових залоз.
Хромофобні ендокриноцити секреторних гранул не мають і слабко фарбуються. Вважають, що вони є малодиференційованими камбіальними клітинами для хромофільних ендокриноцитів. З часом у них формуються секреторні гранули.
Проміжна частина аденогіпофіза сформована меланотропо-
цитами і ліпотропоцитами, секреторні гранули яких фарбуються основними барвниками. Меланотропоцити продукують меланотропін, який впливає на пігментний обмін, а ліпотропоцити – ліпотропін, який стимулює обмін ліпідів.
Ендокриноцити туберальної частини аденогіпофіза мають ку-
бічну форму і їх цитоплазма фарбується основними барвниками. В окремих клітинах є секреторні гранули. Функції ендокриноцитів цієї частини аденогіпофіза до цього часу не встановлені.
Н е й р о г і п о ф і з розвивається з проміжного мозку. Утворений клітинами нейроглії – пітуїцитами і містить накопичувальні тільця, які прилягають до кровоносних капілярів. Накопичувальні тільця – це кінцеві гілочки аксонів нейросекреторних клітин супраоптичних і паравентрикулярних ядер гіпоталамуса, які містять окситоцин і вазопресин. У нейрогіпофізі вони потрапляють у кров. Власні гормони у нейрогіпофізі не синтезуються.
Епіфіз розвивається з дорсальної частини проміжного мозку і мезенхіми. Із нервового зачатка утворюються нейросекреторні клітини і нейроглія, а з мезенхіми – сполучнотканинна строма.
Зовні епіфіз вкритий сполучнотканинною капсулою, від якої відходять перегородки, які ділять його на часточки (рис. 48). Паренхіма часточок утворена нейросекреторними клітинами – пінеалоцитами і гліоцитами.
Пінеалоцити – це крупні відросчасті клітини. Їх відростки біля кровоносних капілярів утворюють булавоподібні розширення, у яких є секреторні гранули і мітохондрії. У цитоплазмі пінеалоцитів добре розвинені синтезуючі органели і лізосоми. Залежно від функціонального стану пінеалоцити поділяють на світлі і темні. Перші мають мало секреторних гранул, а у других їх багато. Гранули фарбуються кислими і основними барвниками. Пінеалоцити продукують серотонін, який в них же перетворюється на гормон мелатонін. Мелатонін забезпечує регуляцію фотоперіодичності роботи органів організму (зміну активності клітин у денний і нічний час). Він також пригнічує діяльність гонадотропних ліберинів гіпоталамуса і гонадотропоцитів аденогіпофіза, гальмуючи передчасне статеве дозрівання, впливає на піг-
170
ментний обмін, процеси поділу і диференціації клітин та виявляє протипухлинну дію. Крім мелатоніну пінеалоцити продукують гормони, які стимулюють діяльність щитоподібної залози, регулюють мінеральний обмін тощо.
Б |
В |
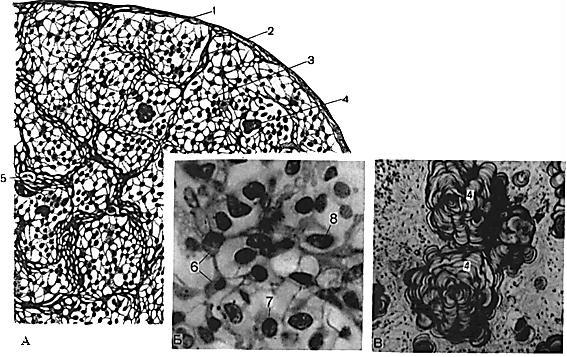
Рис. 48. Мікроскопічна будова епіфіза:
А – схема мікроструктури епіфіза, Б – паренхіма епіфіза, В – мозковий «пісок» в епіфізі; 1 – капсула; 2 – перегородки; 3 – паренхіма; 4 – мозковий «пісок»; 5 – кровоносні судини; 6 – гліоцити; 7 – темні пінеалоцити; 8 – світлі пінеалоцити
Гліоцити – це також відросчасті клітини. Вони виконують переважно опорну функцію відносно до пінеалоцитів. У їх цитоплазмі міститься мало синтезуючих органел і є багато мікрофіламентів. Відростки гліоцитів вплітаються у складові сполучнотканинної строми органа.
Для епіфіза властивий ранній початок вікової інволюції. Вона виявляється атрофією частини пінеалоцитів і розростанням сполучнотканинної строми. В останній утворюється мозковий пісок – накопичення карбонатних і фосфатних солей.
Щитоподібна залоза – це периферичний орган ендокринної системи (рис. 49). Вона побудована зі сполучнотканинної строми і паренхіми. Сполучнотканинна строма утворена пухкою волокнистою сполучною тканиною. У ній є кровоносні й лімфатичні судини та нерви. Сполучнотканинна строма представлена капсулою, яка вкриває орган
171
зовні та трабекулами. Останні відходять від капсули і ділять залозу на часточки, які утворені фолікулами та міжфолікулярними острівцями, між якими є ніжні прошарки пухкої волокнистої сполучної тканини з численними кровоносними судинами.
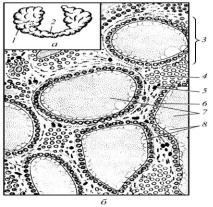
Рис. 49. Щитоподібна залоза великої рогатої худоби (а) та її мікроскопічна будова (б):
1 – частка; 2 – перешийок; 3 – фолікул; 4 – міжфолікулярний острівець; 5 – кровоносний капіляр; 6 – тироцити; 7 – колоїд; 8 – резорбційні вакуолі
Фолікули мають кулясту форму, їх діаметр коливається від 0,02 до 0,7 мм. У них виділяють стінку і порожнину, яка заповнена колоїдом. Останній за помірної функціональної активності залози гомогенний і заповнює усю порожнину фолікулів, при гіперфункції, він має пінистий вигляд і містить вакуолі, а при гіпофункції ущільнюється – між ним і стінкою фолікулів є простори. Стінка фолікулів утворена ендокриноцитами, які називають тироцитами і парафолікулярними клітинами та базальною мембраною.
Тироцити за помірної функціональної активності залози мають кубічну форму, при її гіперфункції вони стають циліндричними, а при гіпофункції і у старих тварин – плоскими. Ядро тироцитів кулясте, а при гіпофункції воно стає видовжено-овальним. У цитоплазмі цих клітин містяться добре розвинені синтезуючі білки органели, лізосоми і є багато секреторних включень. На апікальному полюсі тироцитів є мікроворсинки, а плазмолема базального полюса утворює інвагінації – впинання в цитоплазму. Кількість інвагінацій і мікроворсинок збільшується при посиленні функціональної активності залози. Сусідні тироцити з’єднані десмосомними контактами. Тироцити синтезують йодовмісні гормони тироксин і трийодтиронін, які регулюють окисні процеси, що впливають на всі види обміну речовин в організмі.
Секреторна функція тироцитів відбувається у дві фази: продукування гормонів та їх виведення. У фазу продукування через базаль-
172