Материал: Bovine Viral Diarrhea Virus Diagnosis, Management, and Control

Pathogenesis |
123 |
America were identified as BVDV 2 (Ridpath et al., 1994; Heinz et al., 2000). Further investigation revealed, however, that BVDV 2 encompasses not only strains of high virulence, but also those of moderate to low virulence (Ridpath et al., 2000). Virulence factors associated with BVDV have not yet been defined on an antigenic or genetic level. The occurrence of virulence factors is independent of viral species, although the frequency of virulent strains appears to be higher in BVDV 2 than in BVDV 1. There are regional differences in the relative frequency of BVDV species. Although a high percentage of BVDV strains belong to BVDV 2 in North America (Bolin and Ridpath, 1998; Carman et al., 1998; Fulton et al., 2000), they are infrequently isolated in other countries (Wolfmeyer et al., 1997; Letellier et al., 1999; Sakoda et al., 1999; Tajima et al., 2001; Drew et al., 2002; Vilcek et al., 2002). This is reflected in the different frequency of severe acute BVDV infection in different countries.
Virulence is not correlated with the biotype, since both ncp and cp strains are represented in both BVDV 1 and BVDV 2 and have a wide spectrum of virulence. There are, however, differences between the ncp and cp biotype. ncp BVDV spreads much wider in the host than the homologous cp virus (Spagnuolo-Weaver et al., 1997). The ncp and cp biotype also differ in the way they activate the immune system (Collen and Morrison, 2000). Although ncp BVDV induces a more pronounced humoral response, the response to cp BVDV is skewed toward a cell-mediated immune response. This is particularly evident upon secondary exposure to cp BVDV (Lambot et al., 1997). One can hypothesize that this is mediated by the different effects of ncp and cp BVDV on monocytes and dendritic cells. After in vitro infection, monocyte function is altered by ncp BVDV, but they are killed by cp BVDV. Dendritic cells are not affected by infection with either ncp or cp BVDV (Glew et al., 2003)
The highly variable clinical signs present difficulties for diagnosing BVDV infections in the field. Even if acute BVDV infection is suspected, it may be difficult to confirm, since virus is only transiently detectable (Sandvik et al., 1997). Except for immune status, the role of other host factors on BVDV infection has not yet been determined. However, because of differences in clinical signs and speed of recovery between individual animals after experimental infection, the role of host factors should not be neglected (Liebler-Tenorio et al., 2003a).
VIRUS SPREAD AND DEVELOPMENT OF
LESIONS IN ACUTE BVDV INFECTION
The most frequent route of natural infection is by oronasal uptake of BVDV. The development of clinical signs and distribution and spread of the virus in the host and tissue lesions have been documented in field cases and under experimental conditions (Lambert et al., 1969; Corapi et al., 1990; Wilhelmsen et al., 1990; Traven et al., 1991; Bolin and Ridpath, 1992; Castrucci et al., 1992; Marshall et al., 1996; Spagnuolo-Weaver et al., 1997; Bruschke et al., 1998a; Ellis et al., 1998; Odeon et al., 1999; Archambault et al., 2000; Hamers et al., 2000; Stoffregen et al., 2000; Liebler-Tenorio et al., 2002, 2003a,b). The data were collected primarily for virulent BVDV strains, but some data do exist for strains of low virulence (Wilhelmsen et al., 1990; Traven et al., 1991; Bolin and Ridpath, 1992; Marshall et al., 1996; Bruschke et al., 1998a; LieblerTenorio et al., 2003a). In the following section, the findings for strains of low and high virulence will be described separately.
Strains of low virulence
After intranasal inoculation of calves with BVDV 1 and BVDV 2 strains of low virulence, the animals did not develop overt signs of disease (Wilhelmsen et al., 1990; Traven et al., 1991; Bolin and Ridpath, 1992; Marshall et al., 1996; Bruschke et al., 1998a; Liebler-Tenorio et al., 2003a). The regular monitoring of body temperature showed, however, a mild elevation in temperature for 1–2 days (LieblerTenorio et al., 2003a). This elevation was very shortlived, making it necessary to monitor temperature twice a day.
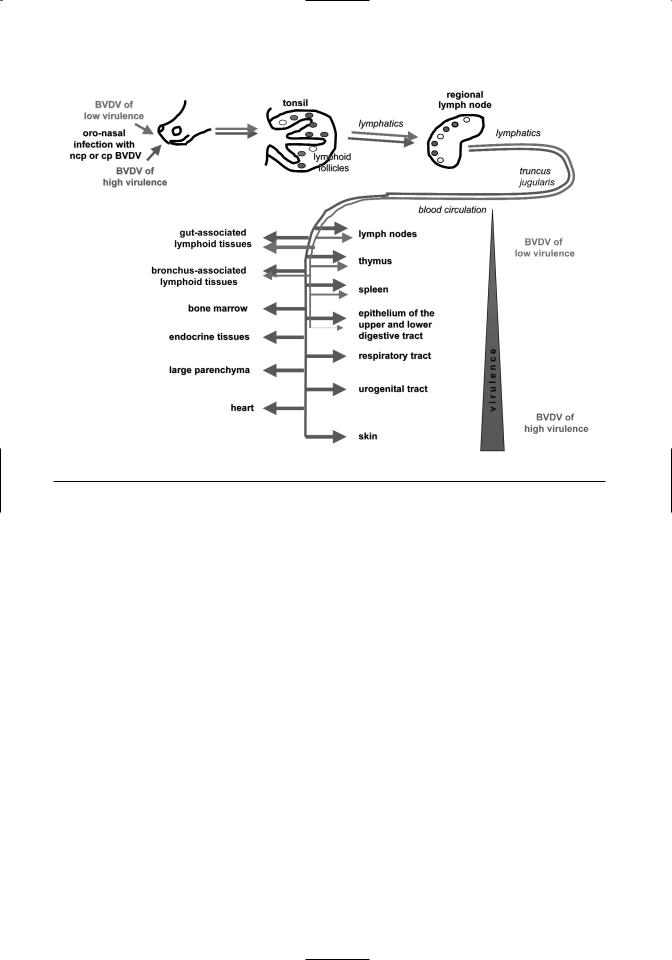
Virus isolation and titration data indicate that BVDV 1 first replicates in the tonsils and nasal mucosa (Bruschke et al., 1998a). Viral antigen was initially found in the tonsils, lymph nodes, and Peyer’s patches and then in spleen and thymus (LieblerTenorio et al., 2003a) (Figure 7.3). Viral antigen was predominantly present within lymphoid follicles and the thymic cortex where it was associated with lymphocytes and stromal cells. Infection in these sites affected the majority of cells present. Multifocal infection of the intestinal mucosa was the only site outside of lymphoid tissues where viral antigen could be demonstrated (refer to Figure 7.3). Viral antigen was not detectable in the bone marrow at any point during infection. Presence of viral antigen was not associated with tissue lesions.
124 |
BVDV: Diagnosis, Management, and Control |
Figure 7.3. Spread of BVDV of low and high virulence in acute BVDV infection.
The widest distribution and the largest amount of viral antigen was found at 6 days postinfection (dpi) followed by a rapid clearance of the virus leading to its disappearance from most sites. The remaining viral antigen was associated with follicular dendritic cells in lymphoid follicles and dendritic cells in the thymus. Viral clearance coincided with the loss of infected lymphocytes leading to a marked depletion of lymphoid follicles and thymic cortex of lymphocytes.
After virus clearance, different degrees of depletion and repopulation were observed in the lymphoid tissues throughout the body. There was a marked difference in the degree of recovery between individual calves indicating the importance of host factors. The reversibility of the lesions was most likely due to the fact that the stromal elements were retained in the lymphoid tissues allowing repopulation with circulating lymphocytes.
Infection with BVDV strains of high virulence
Clinical signs after infection with strains of high virulence are severe but are often nonspecific consisting of high fever, anorexia, depression, and frequently diarrhea (Corapi et al., 1990; Bolin and Rid-
path, 1992; Ellis et al., 1998; Odeon et al., 1999; Archambault et al., 2000; Liebler-Tenorio et al., 2002). Some animals develop severe bleeding (Corapi et al., 1990; Bolin and Ridpath, 1992; Stoffregen et al., 2000). Bleeding is a clinically very dramatic and easy-to-recognize alteration and thus the term hemorrhagic syndrome has been associated with severe acute BVDV infections although it does not occur regularly. The mortality rate may be high even in older cattle (Carman et al., 1998). In experimental infections, early onset of high fever is a consistent finding (Ridpath et al., 2000; Liebler-Tenorio et al., 2002). The other consistent findings are severe progressive lymphopenia and moderate to severe thrombocytopenia (Corapi et al., 1990; Bolin and Ridpath, 1992; Marshall et al., 1996; Ellis et al., 1998; Odeon et al., 1999; Archambault et al., 2000; Hamers et al., 2000; Stoffregen et al., 2000; LieblerTenorio et al., 2002).
The initial spread of highly virulent strains is similar to that of low virulence strains (Liebler-Tenorio et al., 2002, 2003b) (refer to Figure 7.3). There is initial infection of tonsils and lymphoid tissues, but the amount of viral antigen in tissues rapidly exceeds that caused by low virulence strains. The presence of

Pathogenesis |
125 |
highly virulent BVDV is not restricted to follicles in lymphoid tissues but is extended to T-cell–dependent areas. Highly virulent BVDV spreads to the bone marrow. Its presence in myeloid cells and megakaryocytes correlates with a decrease in thrombocyte numbers.
In contrast to BVDV of low virulence, which is cleared from infected tissues, virulent BVDV strains keep spreading (refer to Figure 7.3) resulting in the tissue distribution described for severe acute BVDV (Marshall et al., 1996; Ellis et al., 1998; Odeon et al., 1999; Stoffregen et al., 2000). Antigen of highly virulent BVDV is regularly found in lymphoid tissues, mucosa of the upper and lower digestive tract, the respiratory tract, and endocrine tissues. Eventually, there is hardly any organ that does not contain viral antigen at least in the interstitium. Antigen is often initially present in the interstitium or vascular walls and then in parenchymal cells, indicating a hematogenous spread. Within organs, virus spreads to additional tissue types; e.g., in the intestine where initially the mucosa is antigen-positive, it can later be found in the muscular layers.
Despite the wide distribution of viral antigen, the presence of lesions is restricted. Lesions accompanied by the loss of lymphocytes are consistently observed in lymphoid tissues (Marshall et al., 1996; Ellis et al., 1998; Odeon et al., 1999; Stoffregen et al., 2000; Liebler-Tenorio et al., 2002). Necrosis of epithelium in the upper digestive tract and intestinal crypts may cause erosive to ulcerative lesions along the digestive tract (Marshall et al., 1996; Carman et al., 1998; Ellis et al., 1998; Odeon et al., 1999; Stoffregen et al., 2000).
The discrepancy between presence of viral antigen and lesions is particularly evident in the initial phase of the disease (Liebler-Tenorio et al., 2002). Initially, large numbers of infected cells are present in lymphoid tissues without corresponding morphological lesions. Lesions in lymphoid tissues are delayed. In the thymus, viral antigen is diffusely present in thymic lobules, but lesions are initially multifocal. The same phenomenon can be found in the intestinal mucosa, where viral antigen has a diffuse distribution, but initial lesions can be observed multifocally. In the lungs, frequently foci of acute purulent bronchopneumonia are seen, which do not correlate with the distribution of BVDV antigen in the lung.
Comparison between strains of low and high virulence
Experimental infection with BVDV strains of low and high virulence reveals similar initial infection
and spread but differing amounts of virus in tissues and speed of spreading (Liebler-Tenorio et al., 2003b). This results in a widespread tissue distribution of the virulent strains in later stages of infection, as opposed to an early clearance of the strains of low virulence from tissues. Strains that cause the highest degree of viremia result in the most severe clinical signs (Walz et al., 2001). This indicates that differences in replication, and not interactions between virus and host cell receptors, have the highest impact on the virulence of individual BVDV strains. One can hypothesize that there is a competition between viral spread and reaction of the immune system. The pathogenesis of tissue lesions is unsolved; the discrepancy between the presence of viral antigen and lesions as well as the delayed onset of lesions seen with low and highly virulent BVDV strains might indicate that immune-mediated reactions contribute to the development of lesions.
IMMUNE SUPPRESSION IN ACUTE BVDV
INFECTION
A decrease in the number of both B- and T- lymphocytes in peripheral blood is a consistent finding in acute BVDV infection (Bolin et al., 1985a). The severity of lymphopenia helps discriminate infections with low or highly virulent BVDV strains (Ridpath et al., 2000; Liebler-Tenorio et al., 2003b). Although lymphocyte numbers decrease to more than 60% below the baseline levels, sometimes reaching 90% below the baseline levels after infection with highly virulent strains, the decrease does not exceed more than 50% below the baseline values in strains with low virulence. Lymphopenia correlates well with the infection of and lesions in lymphoid tissue, but it is not resolved yet if lesions are directly virus-induced or if the immune response also contributes to their development.
Experimental infection with virus of low virulence demonstrates clearly that even infections that have a subclinical course and would go unnoticed under field conditions will cause a marked, although transient, immune suppression (Liebler-Tenorio et al., 2003a). This explains reports that combined infections with BVDV have a potentiating effect on several pathogens. Coinfection with BVDV increases the severity of rotaand coronavirus infections in young calves (van Opdenbosch et al., 1981; Kelling et al., 2002; Niskanen et al., 2002b) and the severity of IBRV (infectious bovine rhinotracheitis virus) and BRSV (bovine respiratory syncytial virus) infections (Potgieter et al., 1984; Castrucci et al., 1992; Brodersen and Kelling, 1998, 1999). The

126 |
BVDV: Diagnosis, Management, and Control |
immune response to BRSV is delayed (Elvander et al., 1998). The sequential inoculation of calves with BVDV and Mannheimia haemolytica increased the severity of lung lesions (Potgieter et al., 1984; Ganheim et al., 2003). After vaccination with modified live vaccines for BVDV, there was a decreased response to Mycobacterium paratuberculosis
(Thoen and Waite, 1990). BVDV infection exacerbated the effects of infection with Salmonella sp. (Wray and Roeder, 1987) and led to impaired clearance of bacteremia (Reggiardo and Kaeberle, 1981).
Besides inducing lymphopenia, BVDV has a wide range of effects on the functions of specific as well as innate immune responses (Peterhans et al., 2003). However, immune suppression is not associated with low interferon response or elevated levels of TGF-ß (Charleston et al., 2002). Peripheral lymphocytes are hyporesponsive to mitogens (Muscoplat et al., 1973; Pospisil et al., 1975). Different neutrophil and monocyte functions are impaired (Ketelsen et al., 1979; Roth et al., 1981, Roth and Kaeberle, 1982, 1983; Jensen and Schultz, 1991). Reduction in nonspecific defense has been suggested as the main contribution of BVDV to respiratory tract infections (Potgieter, 1997; Brodersen and Kelling, 1998). A multitude of functional defects are seen in alveolar macrophages from infected calves and those inoculated in vitro with BVDV (Welsh et al., 1995; Liu et al., 1999).
THROMBOCYTOPENIA AND HEMORRHAGES
IN SEVERE ACUTE BVDV INFECTIONS
Thrombocytopenia occurs regularly in cases of severe acute BVD although the reduction of platelets does not always result in marked hemorrhages (Rebhun et al. 1989; Corapi et al., 1990; Bolin and Ridpath, 1992; Ellis et al., 1998; Odeon et al., 1999; Archambault et al., 2000; Hamers et al., 2000; Stoffregen et al., 2000; Liebler-Tenorio et al., 2002). The underlying cause of thrombocytopenia is not completely understood. Necrosis of megakaryocytes, reduced production of thrombocytes by megakaryocytes, increased consumption of thrombocytes in the periphery, and functional defects of thrombocytes have all been suggested as contributing factors (Rebhun et al. 1989; Corapi et al., 1990; Walz et al., 1999, 2001). In some cases of acute BVDV infection, BVDV antigen has been demonstrated in bone marrow (Marshall et al., 1996; Spagnuolo et al., 1997). Experimental infection with BVDV strains of different virulence revealed that not all BVDV strains spread to the bone marrow (Liebler-Tenorio et al., 2003b). The development of
thrombocytopenia is directly related to the infection of bone marrow with BVDV. In the bone marrow, BVDV can be detected in all cellular elements including megakaryocytes (Spagnuolo et al., 1997; Walz et al., 1999; Liebler-Tenorio et al., 2002). Megakaryocytes give rise to thrombocytes by pinching off parts of their cytoplasm. Thus, infection of megakaryocytes appears to be crucial in the development of thrombocytopenia. It is unclear how the infection of megakaryocytes induces a reduction in the number of thrombocytes—e.g., whether there is a reduced production of thrombocytes and/or whether the produced thrombocytes are functionally altered. Investigations by Walz et al. (1999) indicate that both mechanisms are involved.
Bleeding (hemorrhagic diathesis) occurs only when thrombocytes have reached very low numbers. Some infections with highly virulent BVDV may cause mortality early in infection. Since the bone marrow is infected later than other lymphohematopoietic tissues, and thrombocytopenia develops after infection of the bone marrow, the death of the animal may occur before hemorrhagic diathesis becomes established. This might explain the variation in frequency of bleeding observed in the field and in experimental cases of severe acute BVDV infections.
TRANSPLACENTAL/ INTRAUTERINE INFECTIONS
The main economic impact of BVDV infections is caused by intrauterine infections resulting in reproductive dysfunctions (Ross et al., 1986; Kirkbride, 1992; Dubovi, 1994; McGowan and Kirkland, 1995; Moennig and Liess, 1995; Muñoz et al., 1996; Rufenacht et al., 2001). This is due to the fact that BVDV is able to infect the female and male genital tracts, cross the placenta and infect the fetus (refer to Figure 7.2). Transplacental infection may occur in the course of acute infections shortly before or during pregnancy, venereal infection by contaminated semen, and persistent infection (discussed in this section in detail). Even acute infections without clinical evidence of disease may lead to infection of the genital tract and transplacental infection (McGowan et al., 1993). Therefore, any infection shortly before or during pregnancy has severe consequences on reproductive performance.
ROLE OF VIRAL AND HOST FACTORS IN
TRANSPLACENTAL/INTRAUTERINE
INFECTION
The outcome of transplacental/intrauterine infection depends primarily on the time of infection during

Pathogenesis |
127 |
pregnancy and thus the gestational age of the early conceptus or fetus. Infections shortly before or during the first weeks of pregnancy disturb the hormonal balance, cause reduced conception rates, and have direct effect on the early conceptus. In later pregnancy, the outcome of BVDV infection is influenced by the development of target cells, phase of organogenesis, and development of the fetal immune system (Duffel and Harkness, 1985).
The sequelae of infection are also determined by viral properties, especially the biotype of BVDV. Both biotypes are able to infect genital tissues, cross the placenta, and infect the fetus (Scott et al., 1972; Done et al., 1980; Brownlie et al., 1989; Muñoz et al., 1996; Grooms et al., 1998b, 1998c), but the results of infection differ. Infection with the cp biotype appears to cause more damage in the early conceptus than that with ncp BVDV (Vanroose et al., 1998). Only ncp BVDV is able to induce persistent viremia (Brownlie et al., 1989). It has been suggested that the ability to inhibit the induction of type 1 interferon enables the ncp biotype to induce viral persistence (Charleston et al., 2001).
Transplacental infection has been reported for both BVDV 1 and BVDV 2 (Wittum et al., 2001). Dual infection of the fetus with both BVDV 1 and BVDV 2 has also been demonstrated (Brock and Chase, 2000), although differences in the extent of infection were observed between BVDV 1 and BVDV 2. Since the BVDV 2 strain was of higher virulence than the BVDV 1 strain, there was wider tissue distribution of the BVDV 2 strain suggesting an influence of virulence on the efficiency of virus replication.
INFECTION DURING THE PREOVULATORY
PERIOD
Acute infection of cows with BVDV during the preovulatory period may result in viremia at ovulation leading to a decreased conception rate. If conception occurs, the calves are normal, seronegative, and non-viremic at parturition (McGowan et al., 1993). Both ncp and cp BVDV have been demonstrated to infect the ovaries in experimental infections (Grooms et al., 1998b, 1998c). By immunohistochemistry, BVDV can be found in granulosa and stromal cells of ovaries between 8 and 30 days postinfection (Grooms et al., 1998b, McGowan et al., 2003). Histological examination of the ovaries reveal diffuse nonpurulent interstitial inflammation, necrosis of granulosa cells, and necrosis of follicles (Ssentongo et al., 1980; Grooms, et al. 1998c; Fray et al., 2000a; McGowan et al., 2003). Lesions in the
ovaries occur 2–6 days after estrus during the period of seroconversion and may persist for extended time periods (60 days) postinoculation (Ssentongo et al., 1980; Grooms, et al. 1998c, McGowan et al., 2003).
Ovarian lesions are associated with ovarian dysfunctions characterized by decreased secretion of gonadotrophins and sex steroids, particularly progesterone, and absent or reduced preovulatory luteinizing hormone surge (Fray et al., 2000a, 2002; McGowan et al., 2003). The hormonal imbalance leads to decreased growth of ovarian follicles, reduced ovulation rate, reduced numbers of corpora lutea, reduced numbers of collected embryos, and increased numbers of unfertilized ova (Grahn et al., 1984; Grooms et al., 1998a; McGowan et al., 2003). These changes last as long as the lesions persist, at least the first two estrous cycles after infection. This explains the reduced conception rates and the delays in conception (Grooms et al., 1998a).
Ovarian dysfunction is one way that BVDV infections decrease conception rates. Infection of oocytes during infection of ovaries will also impair fertility (Bielanski et al., 1998). The infection of ovaries is multifocal and does not affect all oocytes. Therefore, cows that conceive despite infection produce normal, seronegative, and non-viremic offspring (McGowan et al., 1993). Up to 58% of embryos collected for in vitro fertilization from transiently infected cows were infected with BVDV (Bielanski et al., 1998). Infection of the embryo may result in slightly reduced survival rate and retarded development of the embryo (Archbald et al., 1979; Bielanski and Hare, 1988; Kafi et al., 2002). BVDV can be isolated from retarded embryos (Kirkland et al., 1993).
EXPOSURE OF THE UTERUS TO BVDV AT
INSEMINATION
Exposure of the uterus to BVDV during estrus at insemination causes a decreased conception rate in naive cattle, and seropositive cows have normal conception rates (McClurkin et al., 1979; Whitmore et al., 1981; McGowan et al., 1993; Kirkland et al., 1994). This type of exposure might occur when cows are inseminated with semen contaminated with BVDV or contract acute BVDV infection because of animal movement and mixing around the time of insemination. Experimentally, infection can be reproduced by direct intrauterine instillation of BVDV (Whitemore et al., 1981; Grahn et al., 1984).
Semen contaminated with BVDV may originate from persistently infected bulls, since not all PI bulls are infertile or produce semen of inferior quality