Материал: Bovine Viral Diarrhea Virus Diagnosis, Management, and Control
128 |
BVDV: Diagnosis, Management, and Control |
(Meyling and Jensen, 1988; Wentink et al., 1989; Kirkland et al., 1994). On the other hand, infected semen may also be produced following transient infection of postpuberal bulls (Kirkland et al., 1991; Voges et al., 1998; Niskanen et al., 2002a; Givens et al., 2003). Even after transient infection and clearance of the virus from other tissues and blood, BVDV can persist in semen for months after exposure (Kirkland et al., 1991; Voges et al., 1998; Niskanen et al., 2002a; Givens et al., 2003). The most productive sites of viral replication in the male genital tract are the seminal vesicles and prostate glands causing excretion in the seminal fluid (Kirkland et al., 1991). Insemination with infected semen will cause transient infection of naive females (Kirkland et al., 1994).
The effect of BVDV on gametes and the early conceptus were investigated. In vitro investigations revealed no immediate effect of ncp and cp BVDV on the maturation of oocytes to blastocysts, although BVDV replicated well in cells around the embryo (Kirkland et al., 1996; Tsuboi and Imada, 1996; Kafi et al., 2002). In vitro fertilization of oocytes in the presence of BVDV resulted in reduced fertilization rates (Kafi et al., 2002). There was no uptake of BVDV by embryos after in vitro exposure (Potter et al., 1984). It has been suggested that zona pellucida protects the embryo against infection with BVDV (Singh et al., 1982). After in vitro removal of zona pellucida, 2- and 4-cell embryos were not susceptible to infection with ncp BVDV (Tsuboi and Imada, 1999) and 8-cell embryos were susceptible to cp BVDV (Vanroose et al., 1998). Even extended exposure of embryos with removed zona pellucida had no negative influence on embryonic survival (Bielanski and Hare, 1988). With progressive development, the embryos became increasingly susceptible to infection (Bak et al., 1992, Vanroose et al., 1998; Tsuboi and Imada, 1999).
FETAL INFECTION
After about 30 days of gestation the fetus is susceptible to transplacental infection. There appears to be an increasing efficiency of infection from conception to day 30 postconception (Kirkland et al., 1993). This corresponds with a progressive increase in the loss of pregnancies observed between days 20 and 77 after artificial insemination. Development of placenta with the formation of placentomes, formation of trophoblasts and development of fetal tissues most likely determine the efficiency of infection (Kirkland et al., 1993).
Experimental infections or vaccine trials using
modified live vaccines have shown that fetal infections may result in abortion, persistent infection, stillborn calves, teratogenic effects, and normal calves born with antibodies to BVDV. Although both biotypes of BVDV can infect the fetus, only ncp BVDV is able to establish persistent infection (Brownlie et al., 1989). Infection with cp BVDV results either in abortion or healthy seropositive calves (Brownlie et al., 1989). The fetus, before it becomes fully immune-competent, is affected severely by interauterine BVDV infection. The intrauterine infection of an immune-competent fetus results in a transient infection of the fetus comparable to acute BVD. The fetus is able to mount an immune response and the virus is cleared. Calves are clinically normal and non-viremic. The presence of antibodies to BVDV in serum samples collected before colostrum uptake indicates intrauterine exposure to BVDV. Neutralizing precolostral antibodies have been detected in newborn calves after infections from day 90 to the end of gestation (Kendrick 1971; Orban et al., 1983; Liess et al., 1987).
Abortions
It was recognized early that BVDV may induce abortions (Kahrs and Ward, 1967; Casaro et al., 1971; Kendrick, 1971; Done et al., 1980; Duffell et al., 1984). Abortions occur most frequently in the early stages of gestation (less than 125 days of gestation), although abortions in the late phase of gestation have also been described (Duffel et al., 1984; Roeder et al., 1986). The expulsion of the fetus often occurs not immediately after infection but weeks to months later (Duffel et al., 1984). This also explains the occurrence of mummified fetuses (Scott et al., 1973; Done et al., 1980).
The pathogenesis of abortion is not clear. Most investigations have reported unremarkable, nonspecific lesions of the placenta and placentomes of aborted calves (Casaro et al., 1971; Done et al., 1980; Murray, 1991) but Baszler et al. (1995) found a necrotizing placentitis associated with the presence of viral antigen. It has been speculated that the mild placental lesions might allow opportunistic infections to cross the placenta. BVDV antigen was present in numerous tissues of aborted fetuses, and infiltrates of mononuclear cells were found in several tissues including lung and myocardium (Done et al., 1980; Murray, 1991; Baszler et al., 1995). Therefore, damage to the fetus is considered as important initiator for abortion.
Tissues from pregnant cows infected with an ncp BVDV at 85–86 day of gestation were examined for

Pathogenesis |
129 |
viral antigen by immunohistology (Fredriksen et al., 1999a). BVDV was present in the fetuses at 14 dpi in the lung and liver and at 18–22 dpi in all other tissues examined (lung, liver, spleen, intestine, and brain). The intercotyledonary fetal membranes and the placentomes contained viral antigen at days 18 and 22 postinoculation. BVDV antigen was not detected in maternal tissues or in the placenta between days 7 and 22 postinoculation. This indicates that fetal infection can take place without high concentrations of BVDV in the uterus or placenta of acutely infected cows and that this is a more likely cause of abortion than replication of the virus in the placenta.
Persistent infections
Abortion is not the only outcome of infection in the first trimester of gestation. Infection of the fetus with an ncp BVDV may result in the birth of a PI animal. Generation of PI animals peaks when infection occurs from about 30 until 90 days of gestation and becomes less frequent as the fetus approaches 125 days of gestation (Roeder et al., 1986; Radostits and Littlejohns, 1988; Moennig and Liess, 1995). Persistent infection has been established with both BVDV 1 and BVDV 2. In PI fetuses, BVDV is widely distributed throughout the organs (Fredriksen et al., 1999b). Damage to the fetus is limited and thus pregnancy is maintained. The fetus is immuneincompetent at the time of infection and mounts no reaction to the infecting BVDV. The continued presence of large amounts of virus throughout fetal tissues tolarizes the fetus, when immune competence sets in. Thus the infecting strain of BVDV is not recognized as foreign and there is no immune response to it.
The effect of the persisting BVDV on the fetus and newborn varies (McClurkin et al., 1979; Liess et al., 1984). Persistent fetal infection may result in the birth of normal calves, apparently normal calves with different functional defects or calves with more overt defects. Most have been characterized as “poor-doers.” They may be stillborn or may die within the first few hours or days of life. Surviving calves are often stunted and have growth retardation. In the first year of life, up to 50% higher death rate was found in PI calves when compared to calves not infected (Duffel and Harkness, 1985). Even apparently normal calves have a higher death rate than uninfected calves, because they are predisposed to infections and have a higher risk of severe illness (Barber et al., 1985; Werdin et al., 1989; MuñozZanzi et al., 2003). This is most likely due to functional defects of the immune system causing im-
mune suppression (Roth et al., 1981, 1986). Persistent infection may also cause functional defects of the endocrine system: Diabetes mellitus has repeatedly been reported in PI animals (Taniyama et al., 1995; Buckner, 1997; Murondoti et al., 1999). It has been suggested that different clinical signs associated with persistent infection are predominantly influenced by the time when the fetus becomes infected with earlier infections, resulting more frequently in the birth of normal, healthy virus carriers (Moennig and Liess, 1995).
Teratogenic effects
Congenital defects have been described after infection between days 80 and 150 of gestation. The fetus appears to be the most susceptible to teratogenic effects of BVDV when the immune competence begins to develop, the ability to mount an inflammatory response sets in, and organogenesis is not completed (Duffel and Harkness, 1985).
Intrauterine growth retardation and reduced maturation are observed in numerous tissues, including brain, thymus, muscles, bone, and lung (Done et al., 1980). Lesions in the brain and eyes are most frequent because they are in the final stages of organogenesis during day 100 and 150 of gestation (Duffel and Harkness, 1985). Alterations affecting both organs are referred to as oculocerebellar syndrome (Bielefeldt Ohmann, 1984). The following malformations of the central nervous system have been reported: micrencephaly, cerebellar hypoplasia or aplasia, porencephaly, hydranencephaly, hydrocephalus internus, or dysmyelogenesis (Ward, 1969; Kahrs et al., 1970 a,b; Casaro et al., 1971; Brown et al., 1973, 1974; Scott et al., 1973; Done et al., 1980; Trautwein et al. 1986). Neurological signs of tremor, ataxia, torticollis, or opisthotonus may be seen in newborn calves (Liess et al., 1984). The ocular abnormalities are characterized by reduced pigmentation of the retina, retinal atrophy, optic neuritis, cataract, microphthalmia, and retinal dysplasia (Kahrs et al., 1970b, Brown et al., 1975). Another frequently affected organ is the thymus, which has reduced size at parturition (Done et al., 1980). Other congenital defects are arthrogryposis and alopecia/ hypotrichosis (Casaro et al., 1971; Kendrick, 1971; Bielefeldt-Ohmann, 1984). Frequently BVDV cannot be isolated from or detected in non-PI animals with BVDV-related malformations. In the few cases where BVDV is detected, it is often found locally in the brain or in the cerebrospinal fluid only (Roeder et al., 1986; Trautwein et al., 1987; Liess et al., 1987). Some of the newborns with BVDV-related

130 |
BVDV: Diagnosis, Management, and Control |
malformations are seropositive for BVDV at birth (Liess et al., 1984).
PERSISTENT INFECTIONS
The ability to establish prolonged replication of virus in vivo is important both for the epidemiology and the pathogenesis of BVDV infections. There are two forms of prolonged viral replication. The first form is characterized by viral persistence after intrauterine infection, which results in animals that never clear the virus and thus shed the virus over their entire life span. The second form occurs in some, but not all, animals following acute infection. Such animals shed virus for weeks and months following infection. However, these animals do mount an immune response to the virus and do clear the virus from most tissues over time.
VIRUS PERSISTENCE FOLLOWING
INTRAUTERINE EXPOSURE
The more important form of prolonged viral replication results from intrauterine infection of an im- mune-incompetent fetus because
•PI animals are the main source of virus for establishing acute infections.
•PI mothers are important in establishing PI families by continuous vertical infection.
•Persistent infection is important in the pathogenesis of mucosal disease.
PI animals are characterized by a wide distribution of BVDV throughout their organs, no virusassociated morphological lesions, and no immune response to the persisting BVDV strain. The immune tolerance is selectively restricted to the particular strain of BVDV that has caused the intrauterine infection. Infections with other strains of BVDV will induce an immune response (Fulton et al., 2003). Even single amino acid differences are sufficient for the PI animal to recognize and mount a response to another BVDV strain (Collen et al., 2000). PI animals have normal cellular and humoral responses to other antigens (Houe and Heron, 1993). Recent investigations have centered on immunological mechanisms involved in maintaining viral persistence. It was concluded that tolerance is maintained by nonreactive CD4+ T-lymphocytes, and B-lymphocytes and antigen-presenting cells are normally reactive (Fray et al., 2000b; Glew and Howard, 2001).
scopic lesions (Done et al., 1980; Binkhorst et al., 1983; Roeder et al., 1986). These clinically inconspicuous PI animals are an important source for horizontal and vertical infections. It was demonstrated experimentally that transmission of BVDV to naive animals was much more efficient from PI animals than from those with acute infections (Traven et al., 1991; Niskanen et al., 2000). In PI animals, BVDV is present throughout all organs and tissues (Meyling, 1970; Bielfeldt Ohmann, 1988; Liebler et al., 1991), and thus large scale virus shedding occurs from multiple sites in PI animals. Because BVDV is present in the epidermis; salivary glands; nasal glands; lining of the oral and nasal cavity; and mucosa and accessory glands of the digestive tract, urogenital tract, and mammary glands, the virus can be found in sloughed-off skin cells, oronasal fluids, feces, urine, semen, and colostrum/milk of PI animals. Viral antigen is present in the blood although levels of viremia may change (Brock et al., 1998); thus iatrogenic transmission by contaminated needles or surgical instruments is possible.
Vertical infection
PI cows are able to conceive and give birth to live calves. They have a reduced reproductive performance, which is caused by significant morphological changes in the ovaries leading to reduced follicular maturation (Grooms et al., 1996). Calves born to PI cows are always persistently viremic; this establishes persistent infection in the next generation, thus forming PI families (Straver et al., 1983).
BVDV transmission may occur at the level of oocytes, which can be antigen-positive (Fray et al., 1998). On the other hand, since only about 20% of follicles are positive for BVDV in PI cows (Fray et al., 1998), virus-free calves can be obtained after embryo transfer from PI donors (Wentink et al., 1991; Bak et al., 1992; Smith and Grimmer, 2000). If the oocytes are not infected, infection of the fetus will occur when it becomes susceptible in the early phase of gestation by the continuous presence of BVDV in the uterus, placentomes, and fetal membranes of pregnant PI cows (Fredriksen et al., 1999b). Apparently, pregnancy even enhances the amount of BVDV in the genital tissues (Fredriksen et al., 1999b).
PROLONGED VIRAL SHEDDING FOLLOWING
ACUTE INFECTION
PI animals as source of virus for acute infection
Some PI calves are born without clinical signs of the disease and without any macroscopic or micro-
A prolonged period of viral shed may develop after acute infection of immune-competent animals. Immune-suppressive treatment or stress during in-

Pathogenesis |
131 |
fection may lead to an extended presence of virus in the host for up to 80 days postinoculation (Sandvik et al., 2000). Sometimes virus is retained in certain immunologically privileged sites although a systemic immune response is mounted and virus cleared from most tissues. This type of prolonged viral replication has been described for the male genital system where semen contaminated with BVDV was found for up to 7 months after acute infection (Kirkland et al., 1991; Voges et al., 1998; Niskanen et al., 2002a; Givens et al., 2003).
MUCOSAL DISEASE
Mucosal disease (MD) is the most dramatic clinical evidence of BVDV infection and causes characteristic lesions. The name mucosal disease was coined by Ramsey and Chivers (1953). The authors observed a virus infection in cattle that caused severe erosions, ulcerations, and hemorrhages of the mucosal surfaces of nuzzle, oral cavity, esophagus, forestomachs, abomasum, and the small and large intestines. Further examination revealed severe depletion of spleen, lymph nodes, and thymus. These findings are in accordance with later descriptions by other authors (Schulz, 1959; Bielefeldt Ohmann, 1983; Bolin et al., 1985b; Wilhelmsen et al., 1991). The disease is fatal and the affected animals usually die within 2 weeks after onset of clinical symptoms.
PATHOGENESIS OF MUCOSAL DISEASE
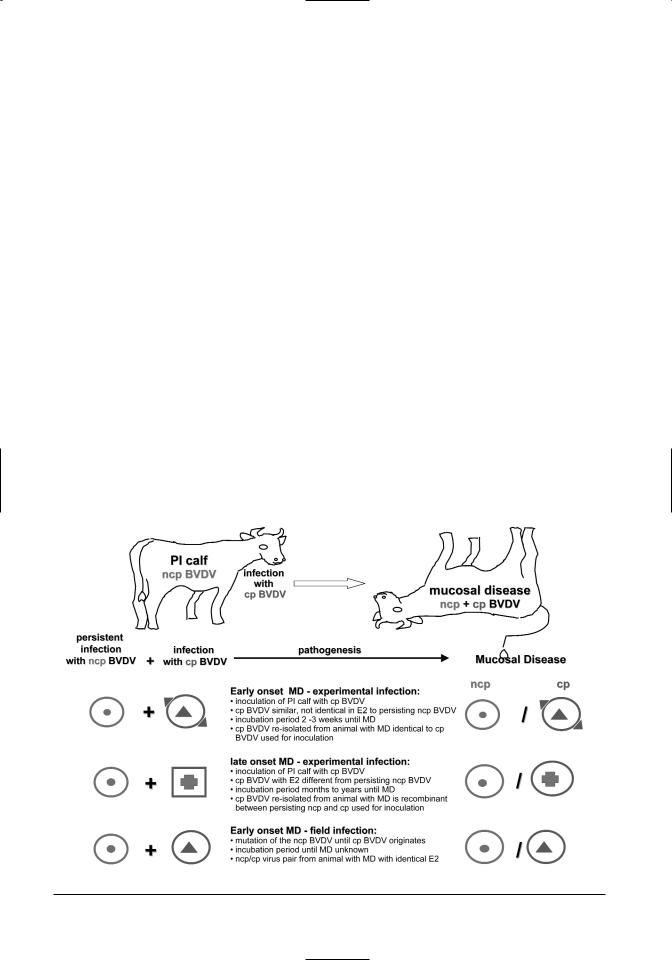
Gillespie et al. (1960) recognized that the virus of mucosal disease was identical with one that causes bovine virus diarrhea (Olafson et al., 1946). Inoculation of immune-competent cattle with BVDVinduced acute BVDV infections and not MD. Although acute BVDV infections infrequently resulted in severe disease with mucosal lesions, they tended to be much more limited and less severe than MD. The pathogenesis of MD remained obscure for several decades. It was only when it was recognized that viral persistence after intrauterine infection of the fetus with ncp BVDV was a prerequisite for the development of MD (Coria and McClurkin, 1978; McClurkin et al., 1984) that researchers were able to induce MD. The first two research teams to induce MD experimentally used slightly different approaches:
•One group inoculated pregnant naive cows during the first trimester of pregnancy with the ncp BVDV of an ncp/cp BVDV pair isolated from an animal with MD. This resulted in the birth of PI calves (McClurkin et al., 1984). When these calves were infected with the cp part of the
BVDV pair, they died from MD (Bolin et al., 1985b).
•The other group identified a herd that contained a number of PI animals. One of these animals developed spontaneous MD. A cp BVDV was isolated from the MD animal. When the remaining PI animals were inoculated with the cp BVDV isolate from the diseased animal, they developed the characteristic signs of MD and died within 2–3 weeks (Brownlie et al., 1984).
Several earlier experiments had revealed that not all experimental infections of PI animals with cp BVDV resulted in MD (Liess et al., 1974; Harkness et al., 1984). The comparison of ncp/cp BVDV pairs from animals with MD showed a very close antigenic relationship between ncp and cp of most virus pairs (Howard et al., 1987). Therefore it was hypothesized that only cp BVDV with close antigenic resemblance (homologous strains) can induce MD (Brownlie and Clarke, 1993; Nakajima et al., 1993). In experimental infections with in vitro selected cp BVDV strains, this could not be confirmed, although close homology appears to favor the development of MD as demonstrated in the following experiments:
•Cytopathic BVDV strains with similar, but not completely identical, reactivity patterns to ncp viruses isolated from PI animals were selected. Reactivity patterns were determined based on binding of monoclonal antibodies produced against the immune dominant surface glycoprotein E2. When the PI animals were inoculated with the selected “homologous” cp BVDV, MD was induced in all animals within 2–3 weeks (Moennig et al., 1990).
•PI animals were inoculated with semihomologous cp BVDV strains. Only some animals succumbed to MD (Bruschke et al., 1998b; Loehr et al., 1998)
•PI animals developed MD after inoculation with an antigenically different cp BVDV (Sentsui et al., 2001).
This led to the hypothesis that in addition to the homology between ncp and cp BVDV, other factors influence the outcome of infection (Bruschke et al., 1998b). Under experimental conditions, MD was reproduced by nasal or intravenous inoculation with cp BVDV. Under field conditions, horizontal infection is possible, especially from animals in the final stages of MD, which shed both biotypes of BVDV. Endogenous development of cp BVDV in PI animals has been discussed as an alternative way of in-
132 |
BVDV: Diagnosis, Management, and Control |
fection (Brownlie and Clarke, 1993; Fricke et al., 2001). When homologous cp BVDV has developed in a PI animal, it will develop MD. During the disease, cp BVDV is excreted and other herd members are infected. Seropositive animals are immune, seronegative animals will develop acute BVD, and other PI herd members infected with the same ncp BVDV will develop MD.
EARLY AND LATE ONSET MUCOSAL
DISEASE
In initial experiments, the first signs of MD developed within 2–3 weeks after inoculation with cp BVDV. However, later reports described the development of MD several months after inoculation with cp BVDV (Brownlie and Clarke, 1993). It was postulated that the course of infection was influenced by the degree of homology between the persisting ncp BVDV and the cp BVDV (Brownlie and Clarke, 1993). Experimental infections of PI animals using cp BVDV strains of variable degrees of antigenic homology as determined by monoclonal antibody reactivity pattern showed that the course of disease is not predictable (Loehr et al., 1998). The following pattern was observed in animals developing MD months to years after inoculation with cp BVDV. An initial, transient acute BVDV infection with the cp
BVDV was followed by a variably long phase without clinical signs and neutralizing antibodies to the cp BVDV. The final phase of infection was denoted by a sudden onset of characteristic signs of MD (Loehr et al., 1998).
Moennig et al. (1993) compared the cp BVDV used for inoculation and the cp BVDV re-isolated from the sick animal from animals that had succumbed to MD months to years after experimental infection. It was found that the re-isolated cp BVDV was a recombination of the cp BVDV used for inoculation with the persisting ncp BVDV. The E2specific monoclonal antibody reactivity pattern of the re-isolated cp BVDV resembled that of the persisting ncp BVDV. However, it expressed the genetic marker of the cp BVDV used for inoculation. The recombinational events were confirmed on a molecular basis by sequencing the E2 and NS2/3 proteins of the viruses involved (Fritzemeier et al., 1995, 1998). The same phenomenon has been described when modified live vaccines containing cp BVDV are used (Ridpath and Bolin, 1995). Such recombinational events may even occur between BVDV 1 and BVDV 2 (Ridpath and Bolin, 1995).
Based on these findings early onset and late onset MD can be distinguished as follows (Figure 7.4): Early onset MD occurs within 2–3 weeks after ex-
Figure 7.4. Pathogenesis of mucosal disease.