Материал: 2_MIKROBIOLOGIYa_temy_5-9_FIZIOLOGIYa_I_IZMENChIVOST_MIKROORGANIZMOV
-образование L- форм микроорганизмов;
-изменение морфологии микроорганизмов;
-изменение тинкториальных свойств;
-изменение ферментативной активности;
-R-S-диссоциации.
Неспецифические реакции возникают в ответ на действие факторов окружающей среды, например, при действии дезинфектантов.
Примерами специфических приспособительных реакций могут быть:
а) синтез новых ферментов (при контакте с антибактериальными препаратами); б) замещение метаболитов антиметаболитами; в) потеря способности метаболита включаться в обменные реакции.
Мутации
Мутации бактерий - это внезапные, ненаправленные изменения первичной структуры ДНК, которые передаются по наследству.
В результате структурной перестройки генов (последовательности нуклеотидов) происходит фенотипическое изменение какого-либо признака или их совокупности (морфологических, биохимических и др.).
Мутации бактерий классифицируют:
1)по протяженности повреждения ДНК:
точечные (повреждения ограничиваются одной парой нуклеотидов),
протяженные, или аберрации (выпадения нескольких пар нуклеотидов - делеции, добавление нуклеотидных пар - дупликации, перемещения фрагментов хромосомы
– транслокации, перестановки нуклеотидных пар – инверсии);
2)по условиям возникновения:
спонтанные (возникают самопроизвольно, без воздействия извне),
индуцированные (при действии внешних факторов — мутагенов);
3)по направленности изменений:
прямые (приводят к потере функции),
обратные, или реверсии (приводят к восстановлению исходного генотипа и свойств);
4)по фенотипическим последствиям:
нейтральные (фенотипически не проявляются какими-либо изменениями признаков),
условно-летальныме (приводят к изменению, но не к утрате функциональной активности фермента и в зависимости от условий окружающей среды микроорганизмы могут сохранять или утрачивать жизнеспособность),
летальные (приводят к полной утратой способности синтезировать жизненно важный фермент или ферменты и гибели бактериальной клетки).
Диссоциация бактерий
S-R-диссоциации возникают при инсертационных мутациях - встраивании в бактериальную хромосому внехромосомных факторов наследственности (R-плазмид, транспозонов или Is-последовательностей, а у возбудителя дифтерии - лизогенных бактериофагов). В результате происходит разделение однородной популяции на два типа колоний:
1)R- колонии (roughнеровный) с неровными краями и шероховатой поверхностью;
2)S- колонии (smoothгладкий) округлой формы и гладкой поверхностью.
Различия между R- и S- формами касаются также антигенных и биохимические свойств бактерий, населяющих колонии, а также их вирулентности, чувствительности к бактериофагам, антимикробным препаратам и других свойств (табл. 15).
Таблица 15
|
Свойства S-R-диссоциантов |
||
S-форма колоний (гладкие) |
|
R-форма колоний (шероховатые) |
|
Равномерное помутнение при |
|
Рост в бульоне в виде осадка |
|
росте в бульоне |
|
|
|
Клетки нормальной морфологии |
|
Клетки измененной формы |
|
|
|
(короткие или длинные, |
|
|
|
кокковидные) |
|
Колонии гладкие, правильной |
|
Колонии шероховатые, с |
|
формы выпуклые |
|
неровными краями, уплотненные |
|
У подвижных видов есть жгутики |
|
У подвижных видов жгутики |
|
|
|
отсутствуют |
|
У капсулообразующих видов |
|
Капсулы отсутствуют |
|
развита капсула |
|
|
|
Биохимически более активны |
|
Биохимически менее активны |
|
Полноценны в антигенном |
|
Неполноценны в антигенном |
|
отношении |
|
отношении |
|
У патогенных видов выражена |
|
Слабо или совсем не вирулентны |
|
вирулентность |
|
|
|
В реакции агглютинации образуют |
|
Мелкозернистый осадок в реакции |
|
рыхлый осадок |
|
агглютинации |
|
обычно выделяются в острый |
|
выделяются чаще при хроническом |
|
период заболевания |
|
течении |
|
Чувствительны к бактериофагу |
|
Менее чувствительны к |
|
|
|
бактериофагу |
|
Менее чувствительны к |
|
Более чувствительны к фагоцитозу |
|
фагоцитозу |
|
|
|
S-формы бактерий более адаптированы к существованию в зараженном организме, |
|||
R-формы – в окружающей среде. |
|||
Причины возникновения диссоциации:
неблагоприятные условия окружающей среды;
старение культуры;
действие иммунных сывороток;
действие бактериофагов и др.
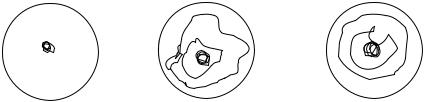
Подавляющее большинство микроорганизмов обладают вирулентностью в S- форме, за исключением возбудителей дифтерии, туберкулеза, чумы, сибирской язвы и некоторых других, проявляющих вирулентность в R-форме:
C.diphtheriae – колонии в виде «маргаритки»;
I.pestis - в виде «кружевного платочка»;
M.tuberculosis - в виде «шагреневой кожи»;
B.anthracis - в виде «локонов» или «львиной гривы».
Рекомбинантная изменчивость
Обмен генетической информацией у бактерий осуществляется путем генетических рекомбинаций.
Генетическая рекомбинация - это передача части хромосомы бактерии-донора бактерии-реципиенту, которая приводит к формированию рекомбинантного генома, сочетающего гены обоих «родителей».
В зависимости от способа передачи различают 3 вида рекомбинаций: трансформации, трансдукции и конъюгации.
Трансформация – это передача генетической информации из разрушенных бактерий-доноров в виде изолированной молекулы ДНК в клетки-реципиенты (опыт Гриффитса, 1928г.)
Этапы трансформации
I этап – адсорбция ДНК донора на рецепторах реципиента. II этап – разрезание двунитевой ДНК эндонуклеазой.
III этап – формирование однонитевых отрезков ДНК. IV этап – синтез ДНК.
V этап – интеграция синтезированного ДНК в геном клетки.
Трансдукция – это передача генетического материала от клетки-донора к клеткереципиенту с помощью умеренных бактериофагов.
Умеренные бактериофаги - вирусы бактерий, обладающие способностью специфически проникать в бактериальные клетки, встраивать свою ДНК в бактериальные хромосомы и реплицироваться в их составе, при этом не вызывая лизиса микроорганизмов. Бактериофаг в таком интегрированном состоянии называют профагом.
Виды трансдукции:
общая – возможен перенос любого гена;
специфическая – переносится строго определенный ген, сцепленный с профагом;
абортивная – переносимый ген интегрирует в геном клетки-донора, и может быть утрачен.
Конъюгация – передача генетического материала от донора к реципиенту при их непосредственном контакте (через цитоплазматический мостик).
Этапы конъюгации
I этап - контакт поверхности клетки-донора и клетки-реципиента.
II этап – прохождение генетического материала через образованный цитоплазматический мостик.
3. Схемы опытов
Опыт модификационной изменчивости
(изучение действия солей желчных кислот на подвижность культуры бактерий Proteus vulgaris.
1 этап. Производят посев P.vulgaris на пластинку среды Плоскирева в центре в виде «бляшки» (1-й пассаж). В качестве контроля исследуемую культуру засевают в виде бляшки на чашку с МПА. Посевы инкубируют при 370С- 18-24 часа.
2 этап. На пластинке МПА обнаруживают рост P.vulgaris не только в виде изолированного скопления («бляшки»), но и «ползучий», «вуалеобразный» рост по всей поверхности среды за счет перемещения бактерий, имеющих жгутики. На среде Плоскирева - рост в виде изолированной бляшки в центре, т.к. под действием солей желчных кислот, содержащихся в этой среде, протей утрачивает жгутики и теряет свою способность передвигаться. С целью определения характера изменчивости производят пересев культуры P.vulgaris 1-го пассажа на пластинку МПА (2-ой пассаж). Посевы инкубируют в термостате при 370С- 18-24 часа.
3 этап. На МПА обнаруживают ползучий рост P.vulgaris
рост P.vulgaris |
рост P.vulgaris |
рост P.vulgaris |
на среде Плоскирева |
на МПА |
на МПА |
(1 пассаж) |
(контроль) |
(2 пассаж) |
Заключение: соли желчных кислот вызывают модификационную (фенотипическую) изменчивость у микроорганизмов вида P.vulgaris, выражающуюся в утрате жгутиков и восстановлении этого признака при устранении неблагоприятного фактора в окружающей среде.
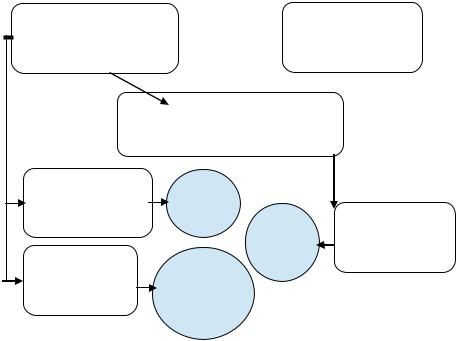
Схема опыта трансформации
(формирование у стрептомициночувствительного штамма B.subtilis резистентности к действию стрептомицина)
Рециптент - B.subtilis Strs |
Донор - B.subtilis Strr |
(стрептомициночувстви- |
(стрептомициноре- |
тельная) |
зистентная) |
Соединение по 1 мл,  инкубация смеси 30 мин при 370С, внесение 1,5 мл раствора ДНК-азы
инкубация смеси 30 мин при 370С, внесение 1,5 мл раствора ДНК-азы
Высев 0,1 мл в |
75 |
|
|
разведении 10-5 |
|
|
|
колоний |
|
|
|
|
|
Высев на МПА со |
|
на пластинку МПА |
|
30 |
|
|
стрептомицином |
||
|
|
||
|
|
колоний |
|
|
|
0,1 мл смеси |
|
Высев на МПА с |
|
|
|
|
|
|
|
добавлением |
|
|
|
стрептомицина |
Отсутствие |
|
|
|
|
|
|
|
роста |
|
|
Частота трансформации рассчитывается как отношение количества клеток-трансформантов к количеству клеток-реципиентов:
30 х 10 |
= 4 х10-6 |
75 х 105 х 10