Материал: Тишин ВБ Новоселов АГ Процессы переноса в технолог аппаратах
a |
1 |
|
d |
R |
2 dT |
qk |
= 0. |
(3.17) |
||
R2 |
|
dR |
|
dR |
|
ρc p |
||||
|
|
|
|
|
|
|||||
В уравнениях (3.16) и (3.17) |
qk является удельной биологиче- |
|||||||||
ской теплотой, выделяемой одной клеткой. |
|
|
|
|||||||
Будем полагать, что температура внутри клетки Tb |
постоянна |
|||||||||
и перенос теплоты из клетки в окружающую среду будет происхо-
дить через клеточную мембрану толщиной |
|
м (см. рис. 3.2), |
внут- |
|||||||||||||||||||||||||||||||||||||||||
ренний радиус которой равен Rb , наружный – R f . |
|
|||||||||||||||||||||||||||||||||||||||||||
|
После первого интегрирования уравнения (3.17) получим |
|
||||||||||||||||||||||||||||||||||||||||||
|
|
R |
2 dT |
|
|
|
|
|
|
|
1 |
|
R |
3 qk |
|
|
|
|
|
C |
, |
|
|
|||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
dR |
|
|
3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
откуда следует |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
dT |
|
|
|
|
|
1 |
|
R |
qk |
|
|
|
|
|
C1 |
|
. |
|
(3.18) |
|||||||||||||||||||
|
|
|
|
|
|
dR |
|
|
3 |
|
|
|
R12 |
|
||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
|
Постоянную интегрирования C1` |
|
находим из следующих усло- |
|||||||||||||||||||||||||||||||||||||||||
вий: R |
Rb , T |
Tb const , dT/dR |
0 : |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||
|
|
|
|
|
|
|
С |
1 |
|
|
|
1 qk |
R |
3 |
. |
|
|
|
|
|
|
(3.19) |
||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3 λ |
|
|
b |
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
После интегрирования уравнения (3.18) получим |
|
||||||||||||||||||||||||||||||||||||||||||
|
|
T |
|
|
|
|
1 |
R |
2 qk |
|
|
|
|
|
|
|
|
C1 |
|
C2 . |
(3.20) |
|||||||||||||||||||||||
|
|
|
|
|
|
6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
R |
|
|
|
|
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
Постоянную интегрирования C2 |
|
найдѐм из следующих усло- |
|||||||||||||||||||||||||||||||||||||||||
вия: R |
R f , T |
T f , где – T f температура на наружной поверхности |
||||||||||||||||||||||||||||||||||||||||||
клетки. Согласно принятым условиям, из уравнения (3.20) следует |
||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
1 |
R2f |
|
|
q |
k |
|
|
|
|
|
1 q |
k |
|
|
R3 |
|
|||||||||||||||||||||
|
|
C2 |
|
T f |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
b |
. |
(3.21) |
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||
|
|
|
|
|
|
|
6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3 |
|
|
|
|
|
|
R f |
|
||||||||
101

Подставив равенство (3.19) и (3.21) в уравнение (3.20), запи-
шем
T T |
|
|
|
1 qk |
(R |
2 |
|
|
R |
2 |
) |
|
|
|
1 qk |
|
R |
3 |
|
1 |
|
1 |
. |
(3.22) |
||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
f |
6 |
|
|
|
|
|
f |
|
|
|
|
|
|
|
|
|
|
|
|
3 |
|
|
|
|
|
|
|
b |
|
R f |
|
R |
|
|
|||||||||||
Из уравнения (3.22) найдѐм разность температур на внутрен- |
||||||||||||||||||||||||||||||||||||||||||||||
ней и наружной поверхностях клетки: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||
T |
|
1 qk |
(R |
2 |
|
R |
2 |
) |
|
|
|
1 qk |
|
|
R |
3 |
|
|
|
1 |
|
|
1 |
|
. |
(3.23) |
||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
k |
|
6 |
|
|
|
|
|
|
f |
|
|
b |
|
|
|
|
|
|
3 |
|
|
|
|
|
|
|
b R |
f |
R |
|
|
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
b |
|
|
|||||
Согласно условию первой задачи, |
|
|
Rb |
|
0 . |
В таком |
случае |
|||||||||||||||||||||||||||||||||||||||
уравнение (3.22) примет вид |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
1 qk |
|
|
|
2 |
|
|
|
|
|
|
|
|
2 |
|
|
|
|
|
|
|
|
|
|
|
(3.24) |
|||||||||
|
|
|
|
T |
|
Tf |
|
|
|
|
|
|
|
|
|
|
|
(R f |
|
|
|
|
|
R |
|
|
). |
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
Разность температур при R |
|
|
|
Rb |
|
|
|
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||
|
|
|
|
|
|
|
|
|
T |
|
|
|
|
1 qk |
R |
2 |
. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
(3.25) |
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
f |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
|
|
|
|
|
k |
|
|
6 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
В уравнениях (3.22)–(3.25) величина qk |
|
есть удельный поток |
||||||||||||||||||||||||||||||||||||||||||||
теплоты, выделяемый одной клеткой. Он зависит от скорости метаболических процессов, протекающих в клетке. Общее количество удельной теплоты, выделяемой всеми находящимися в культиваторе клетками, Ek zqk , где z – количество клеток в единице массы культуральной среды, рассчитываемое по уравнению [22, с. 96]
Ek |
ax b |
dx |
, |
(3.26) |
|
dt |
|||||
|
|
|
|
где x – массовая концентрация биомассы в объѐме культуральной среды Vp . Коэффициент а в данном случае представляет собой коли-
чество теплоты, выделившейся при поддержании жизнедеятельности единицы биомассы в единицу времени, Дж/(кг∙с); коэффициент b –
102

количество теплоты, выделившейся при образовании новых клеток, отнесѐнных к единице биомассы, Дж/кг.
Удельную биологическую теплоту Ek , выделившуюся в единице объема среды в единицу времени, можно назвать ещѐ удельной мощностью биологического источника теплоты Nr /Vp , Дж/(м3∙с).
По своей сути величина Ek имеет тот же смысл, что и диссипация энергии E в равенстве (1.41), т. е. по аналогии с E можно записать, что Ek Nr /Vp . Разница лишь в том, что в равенстве (1.41)
диссипация энергии определяется гидродинамическими условиями в культиваторе, а конкретнее – скоростями сдвига. В уравнении же (3.26) она определяется скоростями метаболических процессов, протекающих внутри живого организма.
Первое слагаемое уравнения (3.26) играет преобладающую роль в стационарной фазе роста, когда основное тепловыделение связано с поддержанием жизнедеятельности клеток и ax >> bdx/dt . Второе слагаемое характеризует тепловыделение в фазе прироста биомассы, когда ax << bdx/dt . При расчѐте теплообмена в культиваторах определяющую роль играет второе слагаемое.
Использование уравнения (3.26) в инженерных расчѐтах затруднено из-за отсутствия надѐжных данных о численных значениях коэффициентов a и b . В литературе иногда приводится суммарное удельное количество биологической теплоты. Например, при культивировании пекарских дрожжей, по данным работы Ю.И. Шишацкого, В.А. Фѐдорова и С.В. Острикова [23], общее количество биологической теплоты, отнесѐнной к единице массы прироста клеток qm , со-
ставляет примерно 4,17 106 |
Дж/кг. Для этого частного случая урав- |
||||
нение (3.26) можно представить в следующем виде: |
|
||||
Ek |
4,17 10 |
6 dx |
. |
(3.27) |
|
|
dt |
||||
|
|
|
|
|
|
Выбор уравнения для вычисления производной в уравнении (3.27) зависит от выбора кинетической модели прироста клеток. Будем считать, что клетки не испытывают дефицита в субстрате, гибель клеток отсутствует, продукты метаболизма постоянно отводятся и потому не влияют на скорость размножения клеток. Если принять
103

экспоненциальный закон развития |
клеток x |
x е k t , то скорость |
|||
|
|
|
|
|
н |
прироста биомассы выразится уравнением |
|
||||
|
dx |
x |
|
e kt , |
(3.28) |
|
|
k |
|||
|
|
||||
|
dt |
н |
|
|
|
|
|
|
|
|
|
где μk – удельная скорость прироста биомассы (клеток), зависящая от многих факторов.
При степенном законе [9] |
|
|
|
||
x x 1 ( t)n |
; |
(3.29) |
|||
|
|
|
н |
|
|
|
dx |
x nynt n 1. |
|
(3.30) |
|
|
|
|
|
||
|
|
|
|
||
|
dt |
н |
|
|
|
|
|
|
|
||
Из уравнения (3.29) следует, что отношение 1/γ представляет собой время удвоения биомассы.
В уравнении (3.29) xн – концентрация биомассы в засевной культуре; γ определяет скорость прироста биомассы и зависит от тех же факторов, что и k , например от концентрации биомассы в на-
чальном засеве, концентрации субстрата в культуральной среде, при аэробном культивировании – от условий аэрации и т. п.
Из уравнений (3.29) и (3.30) получаем выражение для расчѐта удельной мощности источника биологического тепла:
Ek 4,17 106 xнn nt n 1. |
(3.31) |
Уравнение (3.31) устанавливает закон изменения скорости выделения биологического тепла в процессе культивирования. Для случая аэробного культивирования пекарских дрожжей значения 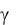 и n могут быть рассчитаны по уравнениям [9, с. 35]:
и n могут быть рассчитаны по уравнениям [9, с. 35]:
a (b |
e c1q ); |
(3.32) |
|||
|
1 |
1 |
|
|
|
n a |
2 |
(b |
e c2q ), |
(3.33) |
|
|
|
2 |
|
|
|
104

где q W /Vp – удельный расход воздуха, здесь W – объѐмный расход воздуха, м3/ч; Vp – объѐм культуральной жидкости в культивато-
ре, м3; a , b и c – коэффициенты, зависящие от начальной концентрации сахара Sн , начального засева хн, температуры, pH среды.
В качестве примера приведим уравнения для расчѐта коэффициентов a , b и c , полученные на основе опытных данных по культивированию хлебопекарных дрожжей при температуре 32 °С
и pH = 4,6:
|
|
a |
|
0,2 |
0,184S |
0,596 |
; |
|
|
|
(3.34) |
||||||||
|
|
1 |
|
|
|
|
|
н |
|
|
|
|
|
|
|
||||
|
b1 |
|
|
|
1 |
|
|
|
|
|
|
|
|
|
|
; |
|
(3.35) |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
0,587 233,7Sн2,514 |
|
|||||||||||||||
|
|
|
|
|
|
|
|
||||||||||||
|
c |
|
|
0,0175 |
16,38S |
2,788 |
; |
|
(3.36) |
||||||||||
|
1 |
|
|
|
|
|
|
|
|
|
|
н |
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
a |
2 |
|
2,033 0,932Sн S 0,037 ; |
|
(3.37) |
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
н |
|
|
|
|
|
|
|
|
|
|
|
|
1 |
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
b |
|
1,386 1,078Sн S 0,179 |
; |
|
(3.38) |
|||||||||||||
|
|
2 |
|
|
|
|
|
|
|
|
|
|
н |
|
|
|
|
|
|
|
|
|
|
|
|
1 |
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
0,0664 |
|
|
||||||||||
c |
2 |
|
0,00293 1,031Sн S |
н |
, |
(3.39) |
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
где Sн – начальная концентрация сахара в массовых долях. Начальный засев xн брался в количестве 8 % от начальной концентрации са-
хара.
Рассмотрим конкретный пример культивирования на мелассном растворе пекарских дрожжей при температуре 32 °С, xн = 2,9 кгАСБ/м3 (АСБ – абсолютно сухая биомасса), Sн = 0,0555, q = 92 1/ч, pH = 4,6. При указанных условиях из уравне-
ний (3.32)–(3.39) γ = 0,285 1/ч , n = 1,346. Подставив имеющиеся дан-
ные в уравнение (3.31) и помня, что произведение nt n 1 имеет размерность 1/ч, получим
|
3 106 t 0,346 |
||
Ek |
|
Вт/м3. |
|
3600 |
|||
|
|
||
105