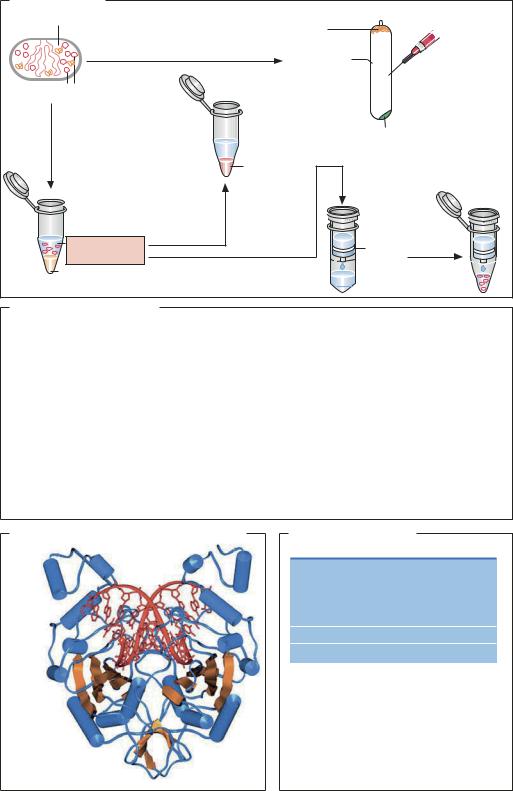
Материал: Шмид Р. Наглядная биотехнология и генетическая инженерия
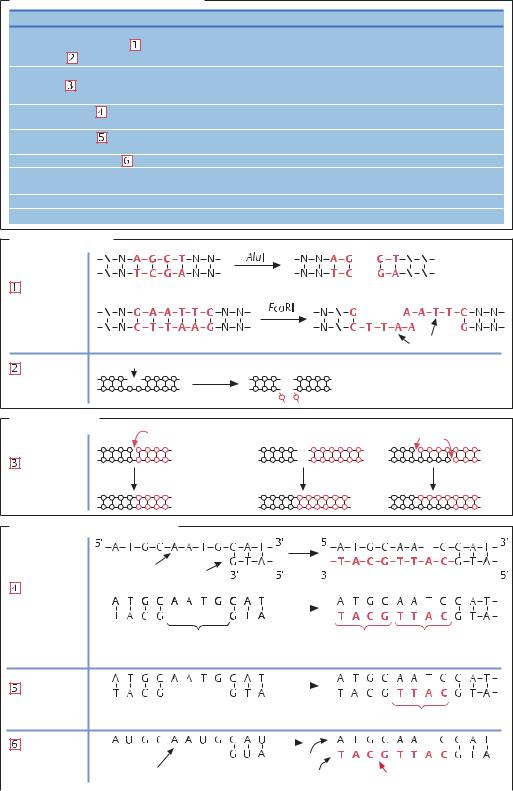
Ферменты, модифицирующие ДНК |
|
|
|
|
Фермент |
|
|
Функция |
|
Расщепление ДНК |
|
|
|
|
Эндонуклеазы рестрикции |
|
|
Расщепляют внутренние фосфодиэфирные связи |
|
Нуклеаза S1 |
|
|
Расщепляет одноцепочечные участки ДНК |
|
Синтез и сшивание ДНК |
|
|
|
|
ДНК-лигаза |
|
|
Восстанавливает разрыв в одной из двух цепей ДНК, |
|
|
|
|
сшивает две молекулы ДНК |
|
ДНК-полимераза I |
|
|
Достраивает вторую цепь на однонитевой матрице, |
|
|
|
|
восстанавливает однонитевые участки |
|
Фрагмент Клёнова |
|
|
Достраивает участки однонитевой ДНК, не обладает |
|
|
|
|
5' → 3'-экзонуклеазной активностью |
|
Обратная транскриптаза |
|
|
Синтезирует ДНК на матрице РНК |
|
Модификации концевых групп |
|
|
|
|
Щелочная фосфатаза |
|
|
Удаляет фосфатные группы на 5'-конце |
|
Полинуклеотидкиназа |
|
|
Вводит фосфатные группы на 5'-конце |
|
Терминальная дезоксинуклеотидил-трансфераза |
Вводит фосфатные группы на 3'-конце |
|||
Расщепление ДНК |
|
|
|
|
N = A, G, C или T |
|
«Тупые» концы |
||
Эндонуклеазы |
|
|
|
|
рестрикции |
|
|
|
|
|
|
|
|
«Липкие» концы |
Разрыв в одной из цепей |
|
|
|
|
Нуклеаза S1 |
|
|
|
|
Сшивание ДНК |
Разрыв в одной |
Лигирование |
Лигирование |
|
|
||||
|
из цепей |
|
«тупых» концов |
«липких» концов |
ДНК-лигаза |
ДНК-лигаза |
или |
ДНК-лигаза |
ДНК-лигаза |
Достраивание ДНК на матрице |
|
|
|
|
|
ДНК-матрица |
Праймер |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Новосинтезированная цепь |
||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
ДНК- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
полимераза I |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Одноцепочечная брешь |
|
|
|
|
|
Предшествующие Брешь закрыта |
||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
нуклеотиды заменены |
||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
(экзонуклеазная активность) |
|||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Фрагмент
Клёнова
Только достройка одноцепочечного участка (отсутствие 5' → 3'-экзонуклеазной активности)
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
РНК |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Обратная |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
транскриптаза |
РНК-матрица |
|
|
|
|
|
ДНК |
|
|
|
Новая цепь ДНК |
|
|||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
233 |
||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
