Материал: Шандра О.А. Нормальна фізіологія. Вибрані лекції _ навч. посіб. _ О.А. Шандра, Н.В. Общіна _ О._ОГМУ, 2005. - 322 с
Отже, можна зазначити, що основною фізіологічною функцією м’язів є рух. Крім того, м’язи беруть участь у теплорегуляції, є місцем відкладання глікогену тощо.
Фізіологічні властивості м’язів: збудливість, провідність, скорочуваність, еластичність (скелетних) і пластичність (гладеньких).
Збудливість чи подразливість — це здатність збудливих тканин
(включаючи і м’язи) відповідати на дію подразника генерацією ПД. Подразливість є властивістю будь-якої живої речовини і виникла разом з нею, але в різних тканинах вона виражена по-різному: в природних умовах збудження м’язів викликається нервовими імпульсами, які надходять із ЦНС. Для того, щоб збудити м’яз в експерименті або при клінічному дослідженні людини, його піддають штучному подразненню електричним струмом. Розрізняють непряме і пряме подразнення м’язів. Непряме подразнення — подразнення м’язів через нерв. Пряме подразнення — це безпосереднє подразнення м’язів. Через те, що збудливість м’язової тканини менша, ніж нервової, при прямому подразненні поріг підвищується. При прямому подразненні струм, поширюючись м’язовою тканиною, діє в першу чергу на закінчення рухових нервів, які знаходяться в ній і збуджують її.
Для доведення прямого подразнення необхідно позбутися нервів. Це здійснюється в такий спосіб: а) кравецький м’яз у верхній 1/4 або 1/5 (проксимальній) частині не має нервових закінчень, тому перерізавши нерв, досягнено дегенерації нервових закінчень, бо без зв’язку з нервовою клітиною відростки нейрона позбавлені живлення (закон Уолера або дегенерації); б) використанням отрути кураре або інших міорелаксантів (наприклад, лістенон, диплацин), які вибірково паралізують рухові пластинки та заважають передачі імпульсів з нерва на м’язи. Мірою збудливості є поріг подразнення.
Провідність— здатність проводити збудження вздовж мембрани м’я- зового волокна. Мірою провідності є швидкість поширення збудження.
Скорочуваність — здатність м’язів скорочуватись або змінювати напруження при збудженні. Залежно від умов, в яких відбувається скорочення, розрізняють три його типи: ізотонічне, ізометричне та змішане.
Ізотонічне — скорочення, при якому не змінюється тонус м’язів, а змінюється їх довжина. Наприклад, підняття м’язом вантажу, який зумовлює його постійне напруження, тобто ізотонічне — скорочення м’яза при постійній напрузі, зафіксований при цьому один кінець м’яза.
Ізометричне — скорочення, при якому змінюється тонус, а довжина залишається без змін, тобто — це таке скорочення, при якому м’яз скоротитися не може, наприклад, якщо обидва кінці його нерухомо закріплені при спробах підняти непосильний вантаж. Природні скорочення м’язів в організмі ніколи не бувають чисто ізотонічними або чисто ізометричними через те, що м’язи, піднімаючи вантаж, укорочуються і разом з тим змінюють своє напруження.
34
Еластичність — властивість м’яза повертатись у початковий стан, якщо його попередньо із нього вивести. М’язи мають невелику, але досконалу еластичність.
Пластичність — здатність гладеньких м’язів утримувати деякий час свою довжину після їх розтягування.
За природою розрізняють такі види скорочень: поодиноке, тетанічне, тонічне і контрактуру.
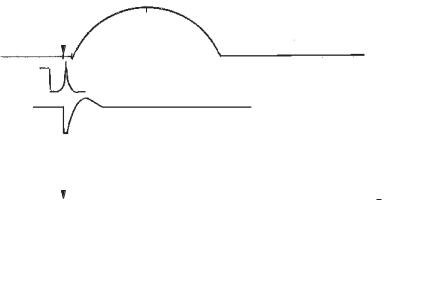
1. Поодиноке скорочення — відповідь м’яза на одиничне подразнення або на один нервовий імпульс. Це елементарна відповідь м’яза, що складається з трьох періодів: латентного, скорочення і розслаблення. Період часу від моменту подразнення до початку скорочення називається прихованим або латентним періодом. Тривалість його в різних м’я- зах різна (скелетний м’яз — 2–3 мс, гладенький м’яз — до 1 с). У цей час у м’язі відбуваються біофізичні та біохімічні процеси, одним із яких є ПД (рис. 8).
А |
|
|
|
|
|
|
|
|
а |
|
|
|
|
|
|
|
|
|
|
в |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
с |
1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||
|
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
-90 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
с |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
в |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
4 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
|
|
|
|
|
|
|
а |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
Б |
|
|
|
|
|
|
5 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
1 |
||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||
0 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
-90 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2 |
||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3 |
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
с |
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
в |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
4 |
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
а |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||||||||||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
Рис. 8. Поодинокі скорочення скелетного (А) і гладенького (Б) м’язів: 1 — крива м’язового скорочення:
а— латентна фаза; в — фаза скорочення; с — фаза розслаблення. 2 — потенціал дії, що виникає в час латентного періоду.
3 — висхідний рівень збудливості м’яза і фази його змін:
а— фаза абсолютної рефрактерності; в — фаза відносної рефрактерності;
с— фаза супернормальної збудливості;
4 — нульовий рівень збудливості
35
Під час поодинокого скорочення m. gastrocnemіus жаби тривалістю в 0,1 с на скорочення йде 0,05 с, на розслаблення — 0,05 с. У однієї і тієї ж тварини різні м’язи мають різну тривалість скорочення залежно від її функції. Вона залежить також від умов (температура, ступінь втомленості). Тривалість скорочення в кожній точці м’язового волокна в десятки разів перевищує тривалість ПД. Тому настає момент, коли ПД пройшов уздовж усього волокна та закінчився (тобто мембрана реполяризувалась), а скорочення охопило все волокно і воно ще продовжує бути укороченим.
Скорочення кожного окремого м’язового волокна при рідкісних поодиноких подразненнях підпорядковуються закону «все або нічого». Це означає, що скорочення, яке виникло як при пороговому, так і при понадпороговому подразненні має максимальну амплітуду. Величина ж поодинокого скорочення всього скелетного м’яза залежить від кількості волокон, що беруть участь у скороченні, а кількість волокон — від сили подразнення. Що сильніший подразник, то більша кількість волокон збуджена. При максимальному скороченні всі волокна м’яза скорочені, тому м’яз підпорядковується «закону градації».
Поодинокі скорочення в природних умовах не зустрічаються, бо до м’язів від спинного мозку відходить «залп» імпульсів, тому навіть дуже швидкі скорочення (рука піаніста) є тетанічними. Такі скорочення переважно штучні.
2. Тетанус. Тривалі скорочення м’яза, викликані частими ритмічними подразненнями, називаються тетанусом. Для одержання тетанусу необхідні часті ритмічні подразнення. Якщо на м’яз до закінчення подразнення подати другий імпульс, то друге скорочення накладається на перше так, що загальне напруження буде більшим, ніж під час першого скорочення, тобто відбувається механічна сумація. Якщо стимули повторюються з короткими інтервалами, то поодинокі скорочення зливаються в тетанус.
Сьогодні не одержав загальноприйнятого пояснення той факт, що напруження, досягнуте під час тетанусу або суперпозиції поодиноких скорочень, більше, ніж сила одного скорочення. Види тетанусу: неповний (зубчастий) і суцільний (гладенький) (рис. 9). При відносно малій частоті подразнень настає зубчастий або неповний тетанус. Наприклад, якщо подразнювати м’яз 5 разів у 1 с, то буде 5 поодиноких скорочень, через те що на одне скорочення необхідно 0,1 с, тимчасом як на 5 скорочень — 0,5 с. Решта 0,5 с — час паузи. Подразнюючи 10 разів у 1 с, одержимо 10 поодиноких скорочень, але без паузи, бо на скорочення пішли всі секунди 0,1×10 = 1 с. Подразнюємо 12 разів у 1 с, кожне повторне скорочення настає до фази розслаблення, бо на одне скорочення використовуватиметься 1:12, 0,08 с, а не 0,1 с. При подразненні 15 разів у 1 с, розслаблення ще більше не буде доходити до 0 лінії.
36
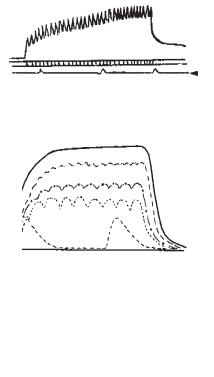
Тобто при зубчастому тетанусі |
|
|
|
|
|
|
є ще елементи поодинокого скоро- |
|
|
1 |
|
||
чення, тому його називають не- |
|
|
|
|||
повним. Так, для м’яза, поодиноке |
|
|
|
|
|
|
скорочення якого дорівнює 0,1 с, |
|
|
ПД |
|||
буде подразнення 20 в 1 с. При |
|
|
|
|
|
|
|
|
t=1 c |
||||
подразненні більше 20 в 1 с настає |
|
|
||||
|
|
|
|
|
||
повний тетанус, тому що при такій |
|
|
|
|
|
|
кількості подразнень м’яз не буде |
|
|
|
|
|
|
встигатирозслаблятися, 1:20=0,05 с |
|
|
|
|
|
|
(весь час, який необхідний для |
|
|
2 |
|
||
скорочення). Важливо те, що при |
|
|
|
|||
|
|
|
|
|
||
тетанусі скорочувальні відповіді |
|
|
|
|
|
|
м’яза підсумовані, електричні ж |
|
|
ПД |
|||
його реакції, тобто ПД не суму- |
|
|
t=1 c |
|||
ються, їх частота відповідає час- |
|
|
||||
тоті ритмічного подразнення, яке |
|
|
|
|
|
|
викликає тетанус. Тобто, якщо |
|
д |
|
|
|
|
дати подразнення 50 в 1 с і спричи- |
, |
|
|
|
||
г |
|
|
|
|||
нити тетанічне скорочення м’яза, |
скороченняСила одиниці.умов |
|
|
|
||
в |
3 |
|
||||
|
|
|||||
то в ньому ми зареєструємо 50 ПД. |
|
|
||||
Це свідчить про те, що тетанічне |
|
б |
|
|
|
|
скорочення складається або супер- |
|
|
|
|
|
|
позується із поодиноких. На це |
|
|
|
|
|
|
вперше вказував ще Гельмгольц |
|
а |
4 |
|
|
|
(1847, теорія суперпозиції). М. Є. |
|
|
|
|
||
|
|
|
|
|
||
Введенський (1885) довів, що хоча |
|
|
|
|
|
|
тетанус складається із поодиноких |
Рис. 9. Види тетанічного скорочен- |
|||||
скорочень, але це не проста сума, |
ня: |
|
|
|
|
|
а значна залежність від частоти |
1 — зубчастий тетанус; 2 — гла- |
|||||
подразнення (а не тільки від сили), |
денький тетанус; |
|
|
|
|
|
а також від фази, на яку припадає |
ПД — потенціали дії, які реєстру- |
|||||
повторне подразнення. Він довів, |
ють при тетанічному скороченні; |
|||||
що при великій частоті подраз- |
3 — зміни амплітуди тетанічних |
|||||
скорочень при збільшенні частоти |
||||||
нення сила скорочення може бути |
||||||
менша, якщо кожне подразнення |
подразнень: |
|
|
|
|
|
припадає на фазу відносної ре- |
а — одинокі скорочення; б, в, г — |
|||||
зубчастий тетанус; д — гладенький |
||||||
фрактерності. До М. Є. Введенсь- |
тетанус |
|
|
|
|
|
кого вважали цю залежність пря- |
|
|
|
|
|
|
мою. М. Є. Введенський увів термін оптимальне (найкраще) «подразнення», яке викликає найбільший ефект. Подразнення, яке за частотою перевищує оптимальне, але викликає менший ефект, називається песимальним (найгірше). Він встановив оптимум і песимум сили та частоти подразнення.
37
Крім поодинокого скорочення, що швидко перебігає, та тетанічного скорочення, що триває довго, деякі м’язи можуть давати тривале скорочення на одноразове подразнення — це тонічне скорочення.
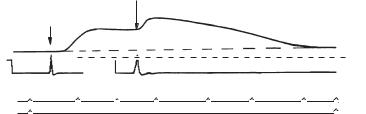
3. Тонічне скорочення. Якщо зареєструвати ПД, буде тільки 1 ПД (рис. 10): це тривале, але не ритмічне скорочення, викликане поодиноким подразненням, називається тонічним скороченням. У тонічному скороченні деякі м’язи перебувають усе життя, наприклад, м’язи кров’яних судин, м’язи ШКТ, м’язи сфінктера сечового міхура. Тонічне скорочення більш характерне для гладеньких м’язів. Скелетні м’язи також можуть давати тонічне скорочення, наприклад, постава людини при сидінні, стоянні. Хоча для скелетних м’язів характерним є тетанічне скорочення, деякі скелетні м’язи перебувають у тонічному скороченні.
Відмінності тетанічного скорочення від тонічного
1.Тетанус — скорочення за природою ритмічне, тонічне — неритмічне. Це підтверджується тим, що при тетанусі відводяться ритмічні ПД, частота яких повністю відповідає частоті ритмічного подразнення (див. рис. 9). Тонічне скорочення ритмічними ПД не супроводжуються. Виникає ПД тільки в момент виникнення або посилення скорочення
(рис. 10).
2.Скорочення тонічне тривале, але слабше від тетанічного. При реєстрації тетанічного скорочення скелетний м’яз підіймає великий (більший) вантаж, ніж при тонічному скороченні, бо при тонічному скороченні м’яз розвиває меншу силу, ніж при тетанічному.
3.Механічна робота при тетанусі супроводжується великою втратою енергії, підвищенням обміну речовин, збільшенням потреби кисню в 10 разів більше, ніж у спокої. Тонічне скорочення супроводжуєть-
6
5
|
0 |
1 |
|
0 |
|
|
|
-70 mV |
-70 mV |
2 |
ПД |
|
3 |
t=7 с |
|
|
|
||
|
|
4 |
t=60 с |
Рис. 10. Крива тонічного скорочення: 1 — скорочення гладенького м’яза; 2 — зареєстровані ПД; 3–4 — відмітка часу; 5, 6 — величина стрілок показує силу подразнень. На більш сильне подразнення (стріка 6) посилюється тонус
38