Материал: Шандра О.А. Нормальна фізіологія. Вибрані лекції _ навч. посіб. _ О.А. Шандра, Н.В. Общіна _ О._ОГМУ, 2005. - 322 с
Трансформація ритму збудження. Здатність нервових центрів змінювати (переробляти) ритм імпульсів, які надходять до них, називається трансформацією ритму. Найчастіше трансформація ритму проявляється в тому, що у відповідь на поодинокий стимул-подразнення нервові центри надсилають до виконавчого органа (ефектора) цілу низку імпульсів, які надходять один за одним з певною частотою. Цим пояснюється й виникнення тетанічного скорочення скелетного м’яза у відповідь на поодиноку стимуляцію відповідного аферентного нерва. Образно кажучи, у відповідь на поодинокий постріл центр відповідає кулеметною чергою. В основі цього явища лежать, головним чином, слідові коливання мембранного потенціалу, особливо тривалість слідового негативного потенціалу. Якщо слідовий негативний потенціал, який супроводжує ПД, великий, то він може досягти критичного рівня деполяризації мембрани і зумовити появу другого імпульсу. У низці випадків трансформація спричинюється збільшенням тривалості ЗПСП, після закінчення першого ПД розвиваються наступні.
Сумація збуджень. Під сумацією збуджень на тілі нейрона розуміють сумацію ЗПСП як місцевого потенціалу. Додавання — процес складання двох або більше однакових реакцій, внаслідок якого результативна реакція буде більшою за своєю величиною, ніж кожна із її складових.
Інтенсивність сумації ЗПСП на тілі нейрона визначає, чи буде цей нейрон збудженим або полегшеним (полегшення — тривале підвищення збудливості нейрона в кірковій речовині великого мозку), а також частоту розрядів збудженого нейрона. Частота розрядів нейрона, тобто частота ПД у серії, яку генерує нейрон, є найважливішою його функціональною характеристикою. Вона дозволяє говорити про інтенсивність збудження конкретного нейрона. Що більша частота розрядів, то сильніше збуджений нейрон.
Розрізняють часову і просторову сумацію збуджень. Якщо ЗПСП швидше (з інтервалом близько 15 мс) надходять аксоном один за одним, то вони додаються в нейроні, досягаючи зрештою порогового рівня деполяризації, потрібного для генерації ПД. Така сумація називається часовою або послідовною. Якщо окремо подразнювати кожний з двох аксонів допороговими подразненнями, то виникають допорогові ЗПСП. Одночасне подразнення аксонів обох сенсорних нейронів зумовлює на мотонейроні сумацію ЗПСП і появу повноцінного ПД, здатного до поширення. Це явище називають просторовою сумацією.
Проте аферентні шляхи, які підходять до нейрона, можуть бути не тільки збуджувальними, а й гальмівними. Імпульси, що надходять цими шляхами, на тілі нейрона перетворюються на гальмівні постсинаптичні потенціали (ГПСП), які сумуються один до одного, так як і до ЗПСП (алгебраїчна дія). Таким чином, у процесі життєдіяльності на тілі кожного центрального нейрона завжди одночасно здійснюється додавання ЗПСП (додавання збуджень) і ГПСП (додавання гальмування). Стан
59
нейрона, незалежно від того, буде він загальмованим чи збудженим (і наскільки), завжди визначається сумацією не лише ЗПСП, а ЗПСП і ГПСП.
Гальмування в центральній нервовій системі
Гальмуваня — нервовий процес, який зумовлюється збудженням і зовнішньо проявляється пригніченням іншого збудження. Воно здатне активно поширюватися нервовою клітиною та її відростками. Започаткував вчення про центральне гальмування І. М. Сєченов (1863), який помітив, що згинальний рефлекс жаби гальмується при хімічному подразненні середнього мозку. Гальмування відіграє важливу роль у діяльності ЦНС, а саме: у координації рефлексів; у поведінці людини і тварин; у регуляції діяльності внутрішніх органів і систем; у здійсненні захисної функції нервових клітин.
Типи гальмування в ЦНС. Центральне гальмування розподіляється за локалізацією на прета постсинаптичне; за характером поляризації (зарядоммембрани) — нагіпертадеполяризаційне; забудовоюгальмівнихнейронних ланцюгів — на реципрокне або поєднане, зворотне та латеральне.
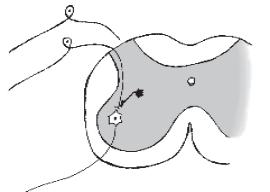
Пресинаптичне гальмування(рис. 13), як свідчить назва, локалізується в пресинаптичних елементах і пов’язане з пригніченням проведення нервових імпульсів у аксональних (пресинаптичних) закінченнях. Гістологічним субстратом такого гальмування є аксональні синапси. До збуджувального аксона підходить вставний гальмівний аксон, який виділяє гальмівний медіатор ГАМК. Цей медіатор діє на постсинаптичну мембрану, що є мембраною збуджувального аксона, і спричинює в ній деполяризацію, яка гальмує вхід Са2+ із синаптичної щілини в закінчення збуджувального аксона і таким чином призводить до пригнічення викидання збуджувальним аксоном медіатора в синаптичну щілину — гальмування реакції. Ще одне пояснення виникнення пресинаптичного гальмування за рахунок іонів Cl-. Механізм цього явища пов’язаний із
Рис. 13. Пресинаптич- 1 не гальмування в спинно-
му мозку:
1 — первинні сенсорні
3нейрони, що містяться в спінальному ганглії поза
спинним мозком; 2 — мо- 2 тонейрон у передніх рогах сірої речовини; 3 — гальмівний нейрон, аксон якого утворює синапс на пресинаптичній терміналі аксона сенсорного нейрона
60
Сl-. З початком розвитку ГПСП Сl- входить у клітину. Коли в клітині стає його більше, ніж зовні, гліцин конформує мембрану і через відкриті її отвори Сl- виходить із клітини. У ній зменшується кількість негативних зарядів, розвивається деполяризація. Такий вид гальмування має назву деполяризаційного.
Пресинаптичне гальмування досягає максимуму через 15–20 мс і триває близько 150 мс, тобто набагато довше, ніж постсинаптичне гальмування. Пресинаптичне гальмування блокується судомними отрутами — бікуліном та пікротоксином, які є конкурентними антагоністами ГАМК.
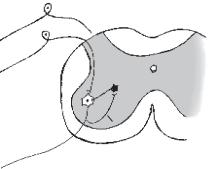
Постсинаптичне гальмування(рис. 14) зумовлене виділенням пресинаптичним закінченням аксона гальмівного медіатора, який знижує або гальмує збудливість мембран соми і дендритів нервової клітини, з якою він контактує. Воно пов’язане з існуванням гальмівних нейронів, аксони яких утворюють на сомі та дендритах клітин нервові закінчення, виділяючи гальмівні медіатори — ГАМК і гліцин. Під впливом цих медіаторів виникає гальмування збуджувальних нейронів. Прикладами гальмівних нейронів є клітини Реншоу в спинному мозку, грушоподібні нейрони (клітини Пуркіньє мозочка), зірчасті клітини кіркової речовини великого мозку та ін. (див. рис. 14). П. Г. Костюк у 1977 р. довів, що постсинаптичне гальмування пов’язане з первинною гіперполяризацією мембрани соми нейрона і виникнення гальмівного постсинаптичного потенціалу (ГПСП), в основі якої лежить збільшення проникності постсинаптичної мембрани для К+. Внаслідок гіперполяризації рівень мембранного потенціалу віддаляється від критичного (порогового) рівня. Тобто відбувається його збільшення — гіперполяризація. Це призводить до гальмування нейрона. Такий вид гальмування називається гіперполяризаційним.
Амплітуда і полярність ГПСП залежать від висхідного рівня мембранного потенціалу самого нейрона.
|
Рис. 14. Пресинаптичне і зворотне |
|
1 |
гальмування: 1 — первинний сенсор- |
|
ний нейрон, аксон якого утворює си- |
||
|
||
|
напс на тілі мотонейрона (2), що |
|
3 |
міститься в передньому розі сірої ре- |
|
човини спинного мозку; 3 — гальмів- |
||
|
ний нейрон, аксон якого утворює си- |
|
2 4 |
напс на тілі мотонейрона (2), і утво- |
|
рює постсинаптичне гальмування. |
Аксон мотонейрона направляє колатераль (4) до гальмівного нейрона і таким чином сам себе гальмує — зворотне гальмування
61
Постсинаптичне гальмування локальне. Розвивається воно градуально, здатне до сумації, не залишає після себе рефрактерності. Є більш оперативним, чітко адресованим та універсальним гальмівним механізмом. За своєю суттю це «центральне гальмування», яке було описане у свій час Ch. S. Sherrington (1906). Залежно від структури гальмівного нейронного ланцюжка, розрізняють такі форми постсинаптичного гальмування: реципрокне, зворотне та латеральне, яке є власне різновидом зворотного.
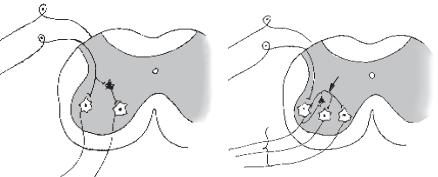
Реципрокне гальмування (рис. 15, а) характеризується тим, що при активізації аферентів збуджуються, наприклад мотонейрони м’язів-зги- начів, та одночасно (з цього боку) гальмуються мотонейрони м’язіврозгиначів, які діють на цей самий суглоб. Відбувається це тому, що аференти від м’язових веретен утворюють збуджувальні синапси на мотонейронах м’язів-агоністів, а через посередництво вставного гальмівного нейрона — гальмівні синапси на мотонейронах м’язів-антагоністів. З фізіологічної точки зору, таке гальмування дуже вигідне, оскільки полегшує рух суглоба «автоматично», без додаткового довільного або недовільного контролю.
Зворотне гальмування (див. рис. 14). У цьому разі від аксонів мотонейрона відходить одна або кілька колатералей, які прямують до вставних гальмівних нейронів, наприклад клітин Реншоу. В свою чергу клітини Реншоу утворюють гальмівні синапси на мотонейроні. У разі збудження мотонейрона активізуються і клітини Реншоу, внаслідок чого відбувається гіперполяризація мембрани мотонейрона і гальмується його діяльність. Що більше збуджується мотонейрон, то відчутніші гальмівні впливи через клітини Реншоу. Таким чином, зворотне постсинаптичне гальмування функціонує за принципом негативного зворотного зв’язку. Є припущення, що цей вид гальмування потрібен для саморегуляції збудження нейронів, а також для запобігання їх перезбудженню та судомним реакціям (див. рис. 15, а).
Латеральне гальмування. Гальмівний ланцюг нейронів впливає не тільки на збуджену клітину, але й на сусідні нейрони, в яких збудження є слабшим або зовсім відсутнє. Таке гальмування називається латеральним, бо ділянка гальмування, яка утворюється, міститься збоку (латерально) від збудженого нейрона. Воно відіграє особливо важливу роль у сенсорних системах, створюючи явище контрасту (рис. 15, б).
Постсинаптичне гальмування переважно легко усувається при введенні стрихніну, який конкурує з гальмівним медіатором (гліцином) на постсинаптичній мембрані. Правцевий токсин також пригнічує постсинаптичне гальмування, порушуючи вивільнення медіатора з гальмівних пресинаптичних закінчень. Тому введення стрихніну або правцевого токсину супроводжується судомами, які виникають внаслідок різкого посилення процесу збудження в ЦНС, зокрема мотонейронів.
62
1
|
1 |
|
|
4 |
5 |
|
|
|
2-МНЗ |
3-МНР |
|
|
|
|
|
3-МНЗ |
2 3 |
|
|
4 |
|
а |
б |
Рис. 15. Реципрокне (а) і латеральне (б) гальмування:
1 — сенсорні первинні нейрони, що містяться в спінальному ганглії поза спинним мозком, його аксон дивергує до мотонейрона-згинача (2-МНЗ) і до гальмівного нейрона (4), аксон якого утворює синапс — (постсинаптичне гальмування) на мотонейроні розгинача (3-МНР); 5 — гальмівний нейрон, аксон якого дивергує до мотонейронів-згиначів (2, 3, 4) і утворює латеральне гальмування. Мотонейрон (2) збуджує гальмівний нейрон за механізмом зворотного гальмування
У зв’язку з розкриттям іонних механізмів постсинаптичного гальмування з’явилася можливість і для пояснення механізму дії брому. Натрію бромід оптимальними дозами широко застосовується в клінічній практиці як седативний (заспокійливий) засіб. Доведено, що такий ефект натрію броміду пов’язаний із посиленням постсинаптичного гальмування в ЦНС.
Роль різних видів центрального гальмування. Головна роль централь-
ного гальмування полягає в тому, щоб у взаємодії з центральним збудженням забезпечувати можливість аналізу і синтезу в ЦНС нервових сигналів, а отже, можливість узгодження всіх функцій організму між собою і з навколишнім середовищем. Цю роль центрального гальмування називають координаційною. Деякі види центрального гальмування виконують не тільки координаційну, а й захисну (охоронну) роль. Припускають, що основна координаційна роль пресинаптичного гальмування полягає у пригніченні в ЦНС малосуттєвих аферентних сигналів. За рахунок прямого постсинаптичного гальмування узгоджується діяльність антагоністичних центрів.
Зворотне гальмування, обмежуючи максимально можливу частоту розрядів мотонейронів спинного мозку, виконує і координаційну роль (узгоджує максимальну частоту розрядів мотонейронів зі швидкістю скорочення м’язових волокон, які вони іннервують), і захисну (запобігає збудженню мотонейронів). У ссавців цей вид гальмування пошире-
63