Материал: Шандра О.А. Нормальна фізіологія. Вибрані лекції _ навч. посіб. _ О.А. Шандра, Н.В. Общіна _ О._ОГМУ, 2005. - 322 с
розглядається як одна із структур гематоенцефалічного бар’єра, який локалізується за межами судинної стінки.
Глія потрібна для синтезу медіаторів ЦНС. Вона відіграє важливу роль і в синаптичній передачі. Нейроглія у свою чергу впливає на гліальні клітини через нейрогліальні синапси, а також за допомогою зміни іонного складу середовища, місцевих струмів та електромагнітного поля, що виникає навколо аксона і до якого глія дуже чутлива. Глія бере участь у білковому і нуклеїново-кислотному забезпеченні нейрона, видаленні з нього продуктів метаболізму. Таким чином, між нейронами та гліальними клітинами існують різноманітні морфофункціональні зв’язки, через які здійснюється взаємокоригуючий вплив. Тому є підстави говорити про те, що до структурно-функціональних одиниць ЦНС належать нейрогліальні комплекси.
Структура і функції центральних синапсів
За допомогою електронної мікроскопії та мікроелектрофізіологічного методу дослідження встановлено, що нейрони не переходять безпосередньо один в одного, а лише контактують між собою через синапси, яких у тисячі разів більше, ніж нервових клітин. Наприклад, кількість синапсів на одній тільки сомі великого мотонейрона спинного мозку може досягати 2000, а на тілі пірамідної клітини кіркової речовини великих півкуль — кількох тисяч. Великі нейрони, як правило, мають більше синапсів. Синапси є також між розгалуженнями аксона і рецепторами або ефекторами (наприклад, м’язові або залозисті клітини).
Розрізняють такі види синапсів: аксосоматичні, аксодендритичні, аксо-аксональні, дендродендритичні. Більшість синаптичних контактів належить до аксодендритичних. Крім того, за функціональними особливостями центральні синапси поділяються на збуджувальні та гальмівні.
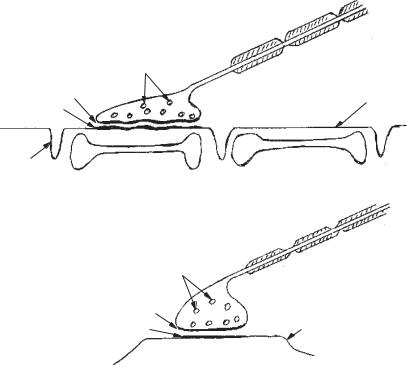
Для відповідних відділів синаптичного сполучення сьогодні прийнято такі терміни: закінчення відростка одного нейрона називають пресинаптичною частиною синапсу (синаптосомою), простір між мембранами, які вкривають пре- і постсинаптичну клітину, — синаптичною щілиною (кілька десятків нанометрів), а ту частину другої клітини, до якої близько підходить закінчення, — постсинаптичною ділянкою. Форма синаптичних сполучень може бути різною — як невеличкі гудзики, «кінцеві ніжки», неправильної форми розширення, що містять сітку нейрофібрил тощо. Найчастішезустрічаєтьсягудзикоподібнийтип(рис. 12, Б).
Для порівняння на рис. 12, А наводиться нервово-м’язовий синапс. Одна з головних функцій пресинаптичного закінчення (синаптосоми) полягає у виділенні медіатора під впливом ПД, що поширюється нервовим волокном. У пресинаптичних закінченнях помітні округлі утворення діаметром 20–50 мкм. Це везикули, або міхурці, які в різній кількості «нагромаджуються» в нервових закінченнях. Везикули розпо-
54
|
|
2 |
А |
6 |
3 |
|
|
|
4 |
|
1 |
5 |
|
|
|
|
|
Са++ |
|
Са++ |
|
|
|
7 |
|
|
|
||
|
|
|
|
8 |
2 |
|
|
|
|
6 |
Б |
4 |
|
|
5 |
1 |
|
Рис. 12. Будова синапсів:
А — периферичний нервово-м’язовий синапс; Б — центральний нервовий синапс.
1 — мембрана клітини (на рис. А — м’язового волокна, на рис. Б — нервової клітини); 2 — нервове волокно; 3 — нервове закінчення — «нервовий ґудзик», «нервова бляшка»; 4 — пресинаптична мембрана; 5 — постсинаптична мембрана; між ними — синаптична щілина; 6 — пухирці медіатора; 7 — поперечні трубочки мембрани м’язового волокна, «Т-система»; 8 — саркоплазматичний ретикулум м’язового волокна — депо Са2+.
діляються по всьому пресинаптичному закінченні, причому їх розподіл і кількість можуть змінюватися залежно від нервової активності. Вони скупчуються біля пресинаптичної мембрани. Везикули містять тисячі молекул хімічного медіатора. Вважають, що везикули лопаються і їхній вміст виливається в синаптичну щілину, здійснюючи «квантове» вивільнення медіатора і перехід збудження з однієї клітини в іншу, як це описано вище. Є дані, що переміщення везикул у бік синаптичної щілини і виділення медіатора відбуваються під впливом Са2+, що входить у нервове закінчення.
55
У відповідь на виділення медіатора з пресинаптичного закінчення та його взаємодію з білковими макромолекулами (хеморецепторами) постсинаптичної мембрани остання змінює проникність для всіх низькомолекулярних іонів, які входять до складу поза- і внутрішньоклітинного середовища. Інактивація нейромедіатора в синаптичній щілині здійснюється шляхом зворотного надходження в пресинаптичне закінчення або ферментативного руйнування. Проникність постсинаптичної мембрани змінюється неоднаковою мірою при збудженні та гальмуванні, внаслідок чого іонні механізми цих двох основних нервових процесів істотно відрізняються.
Крім описаних медіаторних механізмів, встановлено механізми нейромодуляції, які регулюють міжнейронні зв’язки. Нейромодуляторну функцію можуть виконувати нейропептиди та нейрогормони, а також нейромедіатори-супутники, які змінюють спорідненість рецепторів постсинаптичної мембрани до основного медіатора. Нейромодулятори впливають на вивільнення нейромедіатора (пресинаптична модуляція) або на постсинаптичні ефекти медіатора чи на електрогенез постсинаптичного нейрона (постсинаптична модуляція).
Пре- і постсинаптичний модулюючий ефект, як правило, здійснюється вторинними внутрішньоклітинними посередниками. Для пресинаптичної модуляції має значення наявність у пресинаптичних закінченнях рецепторів, які взаємодіють як з власним медіатором, так і з іншими медіаторами (наприклад, з медіатором-супутником).
Медіатори центральної нервової системи
Медіаторами, або нейротрансмітерами, нейронів ЦНС є різні біологічно активні речовини. Залежно від хімічної природи їх можна поділити на 4 групи: 1) аміни (ацетилхолін, норадреналін, дофамін, серотонін); 2) амінокислоти (гліцин, глутамінова, аспарагінова, гамма-амі- номасляна — ГАМК); 3) пуринові та нуклеотиди (АТФ); 4) нейропептиди (речовина Р, вазопресин, опіоїдні пептиди та ін.).
Раніше вважали, що в усіх закінченнях одного нейрона виділяється один медіатор (за принципом Дейла). За останні роки з’ясували, що в багатьох нейронах може міститися 2 медіатори або більше.
За дією медіатори можна поділити на іонотропні та метаболотропні. Іонотропні медіатори після взаємодії з циторецепторами постсинаптичної мембрани змінюють проникність іонних каналів. Метаболотропні медіатори постсинаптичну дію проявляють шляхом активації специфічних ферментів мембрани. Внаслідок цього в мембрані або (найчастіше) у цитоплазмі клітини активуються так звані вторинні посередники (вторинні месенджери), які в свою чергу запускають каскади внутрішньоклітинних процесів, тим самим впливаючи на функції клітин. До основних месенджерів систем внутрішньоклітинної сигналізації зараховують
56
аденілатциклазну та поліфосфоінозитидну. В основі першої лежить аденілатциклазний механізм. Центральною ланкою другої системи є кальціймобілізуючий каскад поліфосфоінозитидів, який контролюється фосфоліпазою С. Фізіологічний ефект цих систем здійснюється шляхом активації специфічних ферментів — протеїнфосфокіназ, кінцевим результатом якого є широкий спектр впливу на білкові субстрати, що можуть піддаватися фосфорилуванню. Внаслідок цього змінюється проникність мембран для іонів, синтезуються і виділяються медіатори, регулюється синтез білків, здійснюється енергетичний обмін тощо. Метаболотропний ефект властивий більшості нейропептидів. Метаболічні зміни, які відбуваються в клітині або на її мембрані під впливом метаболотропних медіаторів, триваліші, ніж при дії іонотропних медіаторів. Вони можуть торкатися навіть геному клітини.
За функціональними властивостями медіатори ЦНС поділяються на збуджувальні, гальмівні та модулюючі. Збуджувальними медіаторами можуть бути різні речовини, які спричинюють деполяризацію постсинаптичної мембрани. Найважливіше значення мають похідні глутамінової кислоти (глутамати), субстанція Р. Деякі центральні нейрони мають холінорецептори, тобто містять на постсинаптичній мембрані рецептори, які реагують із холіновими сполуками, наприклад, ацетилхолін у клітинах Реншоу. Збуджувальними медіаторами можуть бути також моноаміни (норадреналін, дофамін, серотонін). Є підстави вважати, що тип медіатора, який утворюється в синапсі, зумовлений не тільки властивостями закінчення, але й загальним спрямуванням біохімічних процесів у всьому нейроні.
Природа гальмівного медіатора до кінця не встановлена. Вважають, що в синапсах різних нервових структур цю функцію можуть виконувати амінокислоти — гліцин і ГАМК.
Збудження в центральній нервовій системі
Збудження пов’язане з розвитком у нейроні тимчасової деполяризації. Деполяризаційна відповідь нейрона на збудження отримала назву збуджувального постсинаптичного потенціалу (ЗПСП). Внутрішньоклітинний ЗПСП у нейронах ссавців був уперше зареєстрований Д. Екклсом та співробітниками (1966) за допомогою мікроелектродної техніки.
На відміну від збудження, що поширюється (ПД), ЗПСП є місцевим. Воно локальне, градуальне (залежить від сили подразнення, тобто не підпорядковується закону «все або нічого»), здатне до сумації з попередньою деполяризацією, не залишає після себе рефрактерності. За своїми характеристиками ЗПСП є ідентичним потенціалу кінцевої пластинки (ПКП) у нервово-м’язовому сполученні. Проте, якщо ПКП є наслідком активації одного синапсу, то ЗПСП переважно виникає при одночасній активації кількох синапсів.
57
Максимуму ЗПСП досягає протягом 1,5–2 мс, після чого повільно затухає за експонентою. Загальна тривалість ЗПСП багатьох нейронів ЦНС становить близько 15 мс, а амплітуда може досягати 20–30 мВ при величині мембранного потенціалу — 70 мВ. Місцем первинної генерації ПД у нейроні в більшості випадків є початкова частина аксона (аксонний горбик). Висока збудливість цього горбика пояснюється тим, що поріг його деполяризації удвічі менший (15 мВ), ніж усомі нейрона (30 мВ). Це пояснюється тим, що у мембрані аксонного горбика значно більше іонних каналів. Це найбільш збудлива ділянка нейрона (так звана тригерна зона), і лише після її збудження імпульс поширюється, з одного боку, на аксон, а з другого — на сому і дендрити. Потенціал дії нейрона, що реєструється внутрішньоклітинно, має два компоненти: спочатку реєструється потенціал початкового сегмента, або ПС-потенціал, а потім потенціал соми і дендритів — СД-потенціал.
В основі виникнення збудження лежить трансмембранний рух іонів, серед яких домінуюча роль належить Са2+. Збільшення вмісту кальцію в середовищі та цитоплазмі, вплив антагоністів кальцієвої провідності (ВАУК 8644) призводять до збудження, тимчасом як блокатори Са-ка- налів (верапаміл, Д-600) гальмують його розвиток. Є свідчення про можливу участь у генерації ЗПСП Nа+ та Са2+ — збудження нейрона супроводжується змінами метаболізму, зокрема синтезу РНК, та іншими зрушеннями в процесі білкового синтезу, посиленням теплопродукції, поглинанням кисню, які відображають інтенсифікацію життєдіяльності клітини.
Особливості передачі збудження в центральних синапсах
Однобічне проведення. На відміну від нервового волокна, здатного проводити збудження в обох напрямках, у центральному синапсі воно поширюється тільки в одному: від рецептора через вставні нейрони — до ефектора. Це зумовлено тим, що медіатор, вивільнений пресинаптичним закінченням, надходить у синаптичну щілину, діє на рецептори постсинаптичної мембрани, викликаючи там ЗПСП, а потім ПД у постсинаптичній структурі, тобто синапс функціонує як клапан.
Уповільнене проведення збудження. У синапсах ЦНС збудження проводиться повільніше, ніж у нервових волокнах. Це пояснюється особливостями синаптичної передачі (так звана синаптична затримка), а саме дифузією медіатора до постсинаптичної мембрани, виникненням ЗПСП, наростанням його до критичної величини, генерацією ПД. Точні вимірювання дозволили встановити, що для проведення збудження через один синапс потрібно 2–3 мс. Зважаючи на це, можна визначити кількість синапсів у нейронній сітці, через які проходить імпульс у ЦНС.
58