Материал: Прищепа И.Н., Ефременко И.И. Нейрофизиология
32.Период относительной рефрактерности характеризуется: а) повышением возбудимости мембраны клетки; б) понижением возбудимости мембраны клетки;
в) утратой мембраной клетки способности к возбуждению; г) сверхвысокой возбудимостью мембраны клетки.
33.Период абсолютной рефрактерности характеризуется: а) повышением возбудимости мембраны клетки; б) понижением возбудимости мембраны клетки;
в) утратой мембраной клетки способности к возбуждению; г) сверхвысокой возбудимостью мембраны клетки.
34.Период абсолютной рефрактерности совпадает с фазой: а) деполяризации мембраны; б) пика потенциала действия; в) реполяризации мембраны; г) следовых потенциалов.
35.Формулировка закона силы раздражения:
а) чем сильнее раздражение, тем сильнее до определенного предела ответная реакция ткани;
б) пороговая сила раздражителя в определенных пределах находится в обратной зависимости от его длительности;
в) чем больше время действия раздражителя, тем сильнее до определенного предела ответная реакция ткани;
г) пороговая сила раздражителя повышается при уменьшении скорости нарастания стимула.
36.Формулировка закона длительности раздражения:
а) чем сильнее раздражение, тем сильнее до определенного предела ответная реакция ткани;
б) пороговая сила раздражителя в определенных пределах находится в обратной зависимости от его длительности;
в) чем больше время действия раздражителя, тем сильнее до определенного предела ответная реакция ткани;
г) пороговая сила раздражителя повышается при уменьшении скорости нарастания стимула.
37.Формулировка закона градиента (аккомодации):
а) чем сильнее раздражение, тем сильнее до определенного предела ответная реакция ткани;
б) пороговая сила раздражителя в определенных пределах находится в обратной зависимости от его длительности;
в) чем больше время действия раздражителя, тем сильнее до определенного предела ответная реакция ткани;
г) пороговая сила раздражителя повышается при уменьшении скорости нарастания стимула.
38.Формулировка закона силы – времени:
а) чем сильнее раздражение, тем сильнее до определенного предела ответная реакция ткани;
б) пороговая сила раздражителя в определенных пределах находится в обратной зависимости от его длительности;
166
в) чем больше время действия раздражителя, тем сильнее до определенного предела ответная реакция ткани;
г) пороговая сила раздражителя повышается при уменьшении скорости нарастания стимула.
39.На кривой «силы – времени», иллюстрирующей одноименный закон, отмечается реобаза, представляющая собой:
а) силу раздражителя, вдвое большую пороговой величины; б) минимальную силу постоянного тока, способную вызвать воз-
буждение, т.е. порог раздражения; в) наименьшее время, в течение которого должен действовать по-
роговый раздражитель; г) время, в течение которого должен действовать ток, сила кото-
рого вдвое большей пороговой, чтобы вызвать возбуждение.
40.На кривой «силы – времени», иллюстрирующей одноименный закон, отмечается хронаксия, представляющая собой:
а) минимальную силу постоянного тока, способную вызвать возбуждение, т.е. порог раздражения;
б) силу раздражителя, вдвое большую пороговой величины; в) наименьшее время, в течение которого должен действовать по-
роговый раздражитель; г) время, в течение которого должен действовать ток, сила кото-
рого вдвое большей пороговой, чтобы вызвать возбуждение.
Глава 8. ФИЗИОЛОГИЯ СИНАПСОВ. НЕРВНЫЙ ЦЕНТР
Общая физиология синапса
Термины синапс и синаптическая передача были введены в физиологию Ч. Шеррингтоном в 1897 г. Исследуя деятельность ЦНС, он предположил, что между собой нейроны сообщаются с помощью специального синаптического механизма.
Синапс – это морфофункциональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (рис. 26). Все синапсы ЦНС можно классифицировать следующим об-
разом.
По локализации: центральные (головной и спинной мозг) и периферические (нервно-мышечный, нейросекреторный, синапс вегетативной нервной системы). Центральные синапсы можно, в свою очередь, разделить на аксо-аксональные,
167
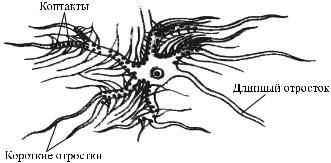
Рис. 26. Расположение контактов на нейроне
аксодендритические, аксо-соматические, дендро-дендритиче-
ские, дендросоматические и т.п.
По развитию в онтогенезе: стабильные (например, синапсы дуг безусловного рефлекса) и динамичные, появляющиеся в процессе индивидуального развития.
По конечному эффекту: тормозные и возбуждающие.
По механизму передачи сигнала: электрические, хими-
ческие, смешанные.
Химические синапсы можно классифицировать:
•по форме контакта – терминальные (колбообразные соединения) и проходящие (варикозные расширения аксона);
•природе медиатора – холинергические (ацетилхолин), адренергические (норадреналин, адреналин), дофаминергические (дофамин), серотонинергические (серотонин), ГАМК-ер- гические (гамма-аминомасляная кислота), глицинергические (глицин), глютаматергические (глютамат), пептидергические (пептиды), пуринергические (АТФ), азотергические (оксид
азота NО) и др.
Электрические синапсы локализованы в стволе мозга. С точки зрения морфологии электрический синапс представляет собой щелевидное образование с ионными мостиками-кана- лами между двумя контактирующими клетками. Петли тока при наличии потенциала действия почти беспрепятственно проходят через такой щелевидный контакт и генерируют ПД в соседней клетке. В целом, синапсы обеспечивают очень быструю передачу возбуждения. Но в то же время такие синапсы обладают двусторонней проводимостью. Кроме того, с их помощью нельзя вызвать торможение эффекторной клетки.
168
Аналогом электрического синапса в гладких мышцах и в
сердечной мышце являются щелевые контакты типа нексуса. Химические синапсы представляют собой окончания ак-
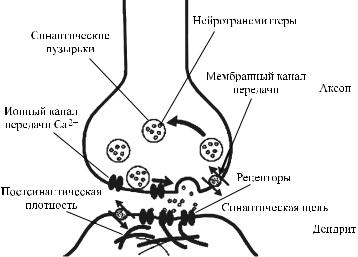
сона (терминальные синапсы) или его варикозную часть (проходящие синапсы). Химический синапс состоит из трех компонентов: пресинаптической части, постсинаптической части и синаптической щели (рис. 27).
Рис. 27. Строение химического синапса
В пресинаптической части содержится медиатор, который под влиянием нервного импульса выделяется в синаптическую щель и, связываясь с рецепторами в постсинаптической части, вызывает ряд физиологических эффектов, в том числе изменение ионной проницаемости постсинаптической мембраны, что приводит к ее деполяризации (в возбуждающих
синапсах) или гиперполяризации (в тормозных синапсах). Пресинаптическая часть представляет собой расширен-
ную конечную часть аксона. В ней содержатся митохондрии, агранулярная эндоплазматическая сеть, нейрофиламенты, нейротрубочки и синаптические пузырьки диаметром 20–
65 нм, в которых находится нейромедиатор. Постсинаптическая часть представлена постсинаптиче-
ской мембраной, содержащей синаптические рецепторы (мем-
169
бранные рецепторы), связывающиеся с нейромедиатором. Мембрана утолщена за счет скопления под ней плотного фи-
ламентозного белкового материала.
Ширина синаптической щели варьирует от 20–30 до 50 нм. Любой химический синапс, независимо от природы медиатора и хеморецептора, активируется под влиянием потенциала действия, распространяющегося к пресинапсу от тела нейрона. Под влиянием потенциала действия происходит деполяризация пресинаптической мембраны, что повышает проницаемость кальциевых каналов пресинаптической мембраны и приводит к увеличению входа в пресинапс ионов Са2+. В ответ на это происходит высвобождение из пресинапса 100–200 порций (квантов) медиатора, что осуществляется путем экзоцитоза. Выйдя в синаптическую щель, медиатор взаимодействует со специфическими рецепторами постсинаптической мембраны. Активированные медиатором рецепторы непосредственно регулируют проницаемость ионных каналов постсинаптической мембраны. Взаимодействие медиатора с постсинаптическими рецепторами изменяет ионную проницаемость. В синапсах, в которых осуществляется возбуждение постсинаптической структуры, обычно происходит повышение проницаемости для ионов Na+ или Са2+, что вызывает деполяризацию постсинаптической мембраны. Эта деполяризация получила название возбуждающего постсинаптического потенциала (ВПСП). Если его величина достигает критического уровня деполяризации, то во внесинаптических областях генерируется ПД. В тормозных синапсах в результате взаимодействия медиатора с рецепторами, наоборот, происходит гиперполяризация (за счет, например, увеличения проницаемости для ионов K+ и Сl–). Этот вид изменения мембранного потенциала получил название тормозного постсинаптического потенциала (ТПСП). В гиперполяризованном состоянии клетка снижает свою возбудимость и благодаря этому прекращает отвечать на внешние раздражители или
уменьшает спонтанную активность.
Одновременно выделившийся в синаптическую щель медиатор может взаимодействовать с рецепторами, расположенными на пресинаптической мембране. Таким способом регулируется интенсивность последующего высвобождения медиатора, т.е. процесс экзоцитоза. Это получило название антидромного эффекта или явления обратной связи.
170