Материал: Прищепа И.Н., Ефременко И.И. Нейрофизиология
ствия в синапсе. Деполяризация этой мембраны соответствует
ВПСП, а гиперполяризация – ТПСП.
Вызванный потенциал – это потенциал действия нейрона, возникающий в ответ на возбуждение рецептора, несущего информацию к этому нейрону.
Мембранный потенциал, или потенциал покоя
Для объяснения причины поляризации мембраны надо рассмотреть свойства самой мембраны, а также ионный состав внутриклеточной жидкости и внеклеточной среды. Мембрана имеет толщину 5–10 нм. Состоит из липидов, белков
имукополисахаридов. Бимолекулярный слой липидов является матриксом мембраны. Белки, вкрапленные в липидный матрикс, образуют каналы для воды и ионов, формируют ионные насосы и т.д. Мукополисахариды, располагаясь в виде «деревьев» на поверхности мембраны, осуществляют рецепторные функции. Мембрана легко проницаема для жирорастворимых веществ, молекулы которых проникают через липидный матрикс. Крупные водорастворимые молекулы, в том числе анионы органических веществ, через мембрану практически не проходят. Кроме того, в мембране нервной клетки существуют каналы, проницаемые для воды, малых ионов
имолекул водорастворимых веществ. В мембране существуют специфические (селективные) натриевые, калиевые, хлорные и кальциевые каналы, т.е. каналы, избирательно пропускающие только натрий, калий, хлор или кальций. Они обладают воротными механизмами и могут находиться в открытом или закрытом состоянии. Помимо этого, мембрана имеет неспецифические каналы для ионной утечки, каждый из которых проницаем для ионов Na+, K+ и Cl– (хотя больше всего проницаем для ионов K+). Каналы не имеют воротных механизмов, всегда открыты и почти не меняют своего состояния при электрических воздействиях на мембрану.
Важным условием существования мембранного потенциа-
ла покоя является отличие ионного состава аксоплазмы от ионного состава внешней среды. Эти различия постоянны и сводятся к тому, что в аксоплазме по сравнению с окружающей средой меньше ионов Na+, больше ионов K+ и значительно больше органических анионов. Что касается катионных раз-
156
личий, то они являются результатом работы так называемого Na+/K+-насоса мембраны, который непосредственно откачивает натрий из клетки в обмен на калий.
В самой мембране мембранный потенциал покоя проявляется как электрическое поле значительной напряженности (105 В/см). Это поле воздействует на макромолекулы мембраны и придает их заряженным группам определенную пространственную ориентацию. Электрическое поле мембранного потенциала покоя обеспечивает закрытое состояние так называемых активационных ворот натриевых каналов и открытое состояние их инактивационных ворот. Этим обеспечивается состояние покоя и готовности к возбуждению.
Потенциал действия
Потенциал действия возникает в ответ на действие порогового раздражителя при достижении критического уровня
деполяризации (рис. 25). Это очень быстрый процесс. В потенциале действия различают пик (спайк) и следовые потен-
циалы. Пик потенциала действия представляет собой кратковременную инверсию внутриклеточного потенциала. Он имеет быструю восходящую фазу и несколько более медленный спад. Общая длительность пика составляет около 3 мс, а ам-
плитуда – 110 мВ, т.е. превышает мембранный потенциал покоя (70 мВ) на 40 мВ. Эту разницу называют овершутом.
Вслед за пиком потенциала действия регистрируются значительно более слабые и длительные отрицательные и положительные следовые потенциалы. Потенциал действия имеет стандартные амплитуду и временные параметры, не зависящие от силы стимула, вызвавшего данный потенциал действия (правило «все или ничего»). Причиной развития потенциала действия является вызываемое критической деполяризацией мембраны открытие ее натриевых и калиевых каналов.
Вход ионов Na+ в клетку обеспечивает восходящую фазу пика потенциала действия, т.е. деполяризацию. В это время происходит инверсия потенциала на мембране. Несколько за-
паздывающий выход ионов K+ участвует в создании нисходящей фазы пика – реполяризации. Быстрый входящий натрие-
вый ток и медленный выходящий калиевый ток возникают практически одновременно, но натриевый развивается, быстрее достигая своего потолка. Если мембранный потенциал
157
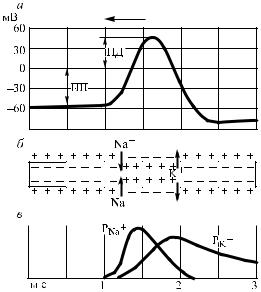
Рис. 25. Схема, иллюстрирующая механизм возникновения потенциала действия (ПД) в нервном волокне:
а – изменения мембранного потенциала; б – схематическое изображение ионных токов; в – изменения проницаемости мембраны для ионов Na+ (PNa+) и K+ (PK+); ПП – потенциал покоя
покоя возвращается к величине, характерной для покоя, то натриевый ток исчезает в 10 раз быстрее, чем калиевый. Можно сказать, что натриевые каналы мембраны и активируются (деполяризацией), и деактивируются (реполяризацией) быстрее, чем калиевые.
Активация каналов определяется открытием их активационных ворот, а деактивация – закрытием этих же ворот.
Вследствие деполяризации, но значительно позже, чем активация, развивается инактивация натриевых каналов, которая
заключается в закрытии специальных инактивационных ворот в натриевых каналах. Инактивация – очень важный механизм, способствующий прекращению пика потенциала действия, развитию временной невозбудимости – рефрактерности. Она устраняется после реполяризации мембраны.
Отрицательный следовой потенциал связан с остаточным током натрия, с накоплением калия в межклеточных щелях. Положительный следовой потенциал обеспечивается остаточ-
158
ным током калия, но главным образом электрогенной работой Na+/K+-насоса мембраны. Работа последнего активируется накоплением ионов Na+ под мембраной, а ионов K+ в межклеточных щелях.
При развитии деполяризации имеют место подпороговые изменения мембранного потенциала, которые проявляются в форме так называемого локального ответа. Первые признаки локального ответа появляются при действии стимула, составляющего 50–75% от пороговой величины. Локальный ответ так же, как и потенциал действия, обусловлен повышением проницаемости мембраны для натрия, но этого повышения недостаточно, чтобы вызвать быструю деполяризацию. Кроме того, она тормозится инактивацией натриевых и активацией калиевых каналов. Амплитуда локального ответа пропорциональна силе подпорогового раздражителя, а не стандартна, как у потенциала действия, т.е. не подчиняется закону «все или ничего». Амплитуда локального ответа увеличивается по мере приближения стимула к пороговой величине, затем он перерастает в потенциал действия, так как скорость увеличения натриевой проницаемости мембраны начинает превышать скорость роста калиевой проницаемости.
В различные фазы развития потенциала действия изменяется возбудимость клеточной мембраны. Во время развития локального ответа возбудимость повышается (мембранный потенциал приближается к критическому уровню деполяризации), во время пика потенциала действия мембрана утрачивает возбудимость, что получило название фазы абсолютной рефрактерности. После окончания пика она постепенно восстанавливается. Этот период называется относительной рефрактерностью и совпадает с периодом реполяризации. Абсолютная рефрактерность объясняется полной инактивацией натриевых каналов и повышением калиевой проводимости. Реполяризация мембраны ведет к реактивации натриевых каналов и снижению калиевой проводимости. Длительность относительной рефрактерности составляет 5–10 мс. При наличии следовой деполяризации она сменяется фазой повышенной возбудимости (супернормальности). Изменение возбудимости клетки имеет значение при ее ответах на ритмическое раздражение. В фазу абсолютной рефрактерности клетка не отвечает на сигнал любой силы. Это имеет защитное значение, так как охраняет клетку от слишком большой частоты раздражителей. Во время абсолютной рефрактерности клетка
159
отвечает только на сверхсильные раздражители, а в фазу супернормальности – на любой, даже подпороговый, раздражитель.
Действие постоянного тока на возбудимые ткани
Генерацию потенциала действия в естественных условиях вызывают местные токи, возникающие между возбужденными и покоящимися участками клеточной мембраны. В связи с этим электрический ток рассматривается как адекватный раздражитель для возбудимых мембран и используется в экспериментах для изучения закономерностей возникновения потенциалов действия. Механизм раздражающего действия тока при всех видах стимулов в принципе одинаков, но наиболее отчетливо он выявляется при использовании постоянного тока.
Полярный закон раздражения заключается в следующем: при раздражении нерва или мышцы постоянным током возбуждение возникает в момент замыкания постоянного тока только под катодом, а в момент размыкания – только под анодом. Этот закон сформулирован немецким физиологом Э. Пфлюгером в 1859 г. Доказывается он следующим образом: умерщвляют участок нерва под одним из электродов, а второй электрод ставят на неповрежденный участок. Если с неповрежденным участком связан катод, то возбуждение возникает в момент замыкания тока, а если анод – то при размыкании. Порог раздражения при размыкании, когда возбуждение возникает под анодом, значительно выше, чем при замыкании, когда возбуждение возникает под катодом. Это объясняется тем, что при приложении к ткани анода происходит гиперполяризация мембраны, а при приложении катода – ее деполяризация. Потенциал действия возникает в тот момент, когда деполяризация мембраны достигает критического уровня. Этот критический уровень деполяризации не зависит от характера применяемого стимула, а определяется только свойствами мембраны.
Скорость, с которой происходит деполяризация, зависит также от силы раздражающего тока. При слабой силе тока деполяризация развивается медленно, поэтому для возникновения потенциала действия стимул должен быть более длительным. В случае раздражающего тока большей силы растет
160