Материал: Прищепа И.Н., Ефременко И.И. Нейрофизиология
Для некоторых нейронов активное состояние возникает спонтанно, автоматически, причем чаще всего автоматия нейрона проявляется периодической генерацией серии импульсов. Примером таких нейронов-пейсмекеров (водителей ритма) являются нейроны дыхательного центра продолговатого мозга. Иногда такие нейроны называют фоновоактивными. По характеру реакции на приходящие импульсы они делятся на тормозные и возбуждающие. Тормозные нейроны урежают свою фоновую частоту разрядов в ответ на внешний сигнал, а возбуждающие увеличивают частоту фоновой активности.
Существует три вида фоновой активности нейронов – не- прерывно-аритмичный, пачечный и групповой. Непрерывноаритмичный вид активности проявляется в том, что фоновоактивные нейроны генерируют импульсы непрерывно с некоторым замедлением или увеличением частоты разрядов. Такие нейроны обычно обеспечивают тонус нервных центров. Фоновоактивные нейроны имеют большое значение в поддержании уровня возбуждения коры и других структур мозга. Число фоновоактивных нейронов увеличивается в состоянии бодрствования.
Групповая форма активности характеризуется периодическим появлением группы импульсов, сменяющихся периодом молчания.
Состояние торможения проявляется в том, что фоновоактивный нейрон или нейрон, получающий возбуждающее воздействие извне, прекращает свою импульсную активность. В основе торможения лежит явление гиперполяризации нейрона (характерно для постсинаптического торможения) или активное прекращение поступающей импульсации от других нейронов, что наблюдается в условиях пресинаптического торможения.
Физиология нервных волокон
Главной функцией нервных волокон является проведение нервных импульсов, в возникновении которых основную роль играет плазматическая мембрана, обладающая высокой возбудимостью. Механизм проведения нервного импульса по миелинизированным и немиелинизированным волокнам различен.
Распространение нервного импульса по немиелинизированным волокнам связано с появлением местных круговых
131
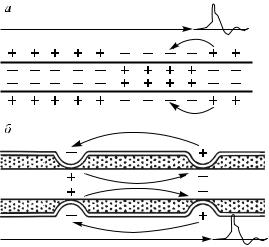
токов, которые возникают при прохождении через мембрану аксона ионов калия, натрия, кальция. Перемещение различно заряженных ионов из аксона в окружающую аксон жидкость приводит к возникновению разности потенциалов между внутриаксональной и наружной средами. Появившаяся разность потенциалов возбуждает участок аксона. В нем также возникают ионные токи, устанавливается разность потенциалов, которая возбуждает следующий участок, и так далее по аксону до синапса (рис. 16, а).
Скорость распространения возбуждения по волокну определяется его диаметром – чем больше диаметр, тем выше скорость проведения возбуждения.
В миелинизированных волокнах скорость проведения нервных импульсов значительно выше, чем в немиелинизированных, хотя и в этих волокнах сохраняется прямая пропорциональная зависимость скорости проведения возбуждения от его диаметра. В миелинизированных волокнах круговые электрические токи возникают только в области перехватов Ранвье. Именно здесь количество натриевых каналов достигает 12 тыс. на 1 мкм2, что значительно больше, чем в любом другом участке волокна. В результате эти участки являются наиболее возбудимыми, и поэтому именно в них генерируется
Рис. 16. Проведение возбуждения в немиелинизированных (а) и миелинизированных (б) нервных волокнах
132
очередной потенциал действия. Он, в свою очередь, вызывает генерацию потенциала действия в соседнем перехвате Ранвье, а возникший в этом перехвате новый потенциал действия вызывает возбуждение следующего перехвата Ранвье. В целом, все это приводит к скачкообразному, или сальтаторному, проведению возбуждения по нерву (рис. 16, б). Такой механизм обеспечивает гораздо более быстрое (примерно в 50 раз) распространение потенциала действия, чем в немиелинизированных волокнах. При этом скорость проведения возбуждения по миелинизированному волокну прямо пропорциональна и диаметру волокна, и длине между перехватами Ранвье, т.е. чем больше диаметр и чем длиннее интервалы между пере-
хватами Ранвье, тем выше скорость проведения возбуждения. Нервное волокно обладает возбудимостью и лабиль-
ностью. Возбудимость миелинизированных нервных волокон выше, чем немиелинизированных. Кроме того, у миелинизированных волокон более высокая лабильность по сравнению со всеми другими нервными образованиями, включая и немиелинизированные нервные волокна. Например, известно, что миелинизированные волокна могут воспроизводить до 1000
импульсов в 1 с.
Законы проведения возбуждения по нервным волокнам. Выделяют следующие законы проведения возбуждения по нервным волокнам: 1) закон изолированного проведения возбуждения; 2) закон анатомической и физиологической целостности нервного волокна; 3) закон двустороннего проведения возбуждения по нервному волокну; 4) закон практической неутомляемости нервных волокон; 5) закон прямо пропорциональной зависимости скорости проведения импульса от диа-
метра нервного волокна.
Закон изолированного проведения возбуждения утверждает, что проведение возбуждения по отдельным нервным волокнам, проходящим в составе нерва, происходит изолированно, независимо от других волокон. Возможность изолированного проведения возбуждения имеет большое физиологическое значение, так как обеспечивает, например, изолированность сокра-
щения каждой нейромоторной единицы.
Закон анатомической и физиологической целостности нервного волокна утверждает, что необходимым условием проведения возбуждения в нерве является не только его анато-
133
мическая непрерывность, но и физиологическая целостность. Если нарушить свойства мембраны волокна (перевязка, блокада новокаином, аммиаком и др.), то проведение возбужде-
ния по волокну прекращается.
Закон двустороннего проведения возбуждения по нервному волокну утверждает, что любое нервное волокно способно проводить возбуждение в обоих направлениях. В этом можно убедиться, если наносить искусственное раздражение на волокно – потенциалы действия будут распространяться в обе
стороны от места раздражения.
Закон практической неутомляемости нервных волокон, который был сформулирован Н.Е. Введенским, указывает на то, что нервное волокно обладает малой утомляемостью. Действительно, проведение возбуждения по нервному волокну не нарушается в течение длительного (многочасового) эксперимента. Считают, что нервное волокно относительно неутомляемо вследствие того, что процессы ресинтеза энергии в нем идут с достаточно большой скоростью и успевают восстановить траты энергии, происходящие при прохождении возбуждения.
В момент возбуждения энергия нервного волокна тратится на работу Na+/K+-насоса. Особенно большие траты энергии происходят в перехватах Ранвье вследствие значительной
плотности здесь Na+/K+-насосов.
Закон прямо пропорциональной зависимости скорости проведения импульса от диаметра нервного волокна был
установлен лауреатами Нобелевской премии американскими физиологами Дж. Эрлангером и Г. Гассером (1944). На основании этого закона авторы предложили классификацию нервных волокон. Они разделили все нервные волокна в зависимости от таких показателей, как диаметр, возбудимость, временные характеристики потенциала действия и его компонентов, а также скорость проведения возбуждения, на три основные группы – А, В и С (табл. 1).
Группа А – это наиболее толстые миелинизированные моторные и чувствительные волокна; группа В – это миелинизированные преганглионарные волокна автономной нервной системы с низким содержанием в них миелина; группа С – это немиелинизированные постганглионарные волокна симпатической нервной системы. Группа А неоднородна – в ней выделены три основные подгруппы (α, β, γ).
134
Таблица 1. Свойства различных нервных волокон млекопитающих
Тип |
Диа- |
Ско- |
Длитель- |
|
|
ность по- |
|
||||
рость |
|
||||
во- |
метр, |
тенциала |
Функции |
||
проведе- |
|||||
локна |
мкм |
действия, |
|
||
|
|
ния, м/с |
мс |
|
|
|
|
|
|
||
|
|
|
|
|
|
А-α |
13–22 |
70–120 |
0,4–0,5 |
Эфферентные волокна, проводящие |
|
|
|
|
|
возбуждение к скелетным мышцам, |
|
|
|
|
|
афферентные волокна, проводящие |
|
|
|
|
|
возбуждение от мышечных рецеп- |
|
|
|
|
|
торов |
|
А-β |
8–13 |
40–70 |
0,4–0,6 |
Афферентные волокна, проводящие |
|
|
|
|
|
возбуждение от рецепторов прикос- |
|
|
|
|
|
новения и сухожильных рецепторов |
|
А-γ |
4–8 |
15–40 |
0,5–0,7 |
Афферентные волокна, проводящие |
|
|
|
|
|
возбуждение от рецепторов прикос- |
|
|
|
|
|
новения и давления; эфферентные |
|
|
|
|
|
волокна – к мышечным веретенам |
|
В |
1–3 |
3–14 |
1,2 |
Преганглионарные волокна вегета- |
|
|
|
|
|
тивной нервной системы |
|
С |
0,5–1,0 |
0,5–2,0 |
2,0 |
Постганглионарные волокна вегета- |
|
|
|
|
|
тивной нервной системы; аффе- |
|
|
|
|
|
рентные волокна, проводящие воз- |
|
|
|
|
|
буждение от рецепторов боли, дав- |
|
|
|
|
|
ления и тепла |
Объединение нейронов как один из принципов организации работы мозга
Для различных структур мозга характерны определенные типы нейронной организации. Нейроны, выполняющие одну и ту же функцию, образуют группы, популяции, ансамбли, колонки, ядра. В коре большого мозга, мозжечке нейроны формируют слои клеток. Каждый слой имеет свою специфиче-
скую функцию.
Серое вещество мозга. Скопления нейронов и нейроглии образуют серое вещество мозга, которое неоднородно. В нем имеются участки концентрации нейронов, где их тела очень плотно располагаются относительно друг друга, а также области, где концентрация нейронов невысокая. Области высокой концентрации нейронов получили название ядер серого ве-
135