Материал: Национальная+программа+Недостаточность+витамина+Д+2018
relationship of physical activity to bone mineral accrual in |
нительной ткани, особенно ревматоидным артритом [6–8]. |
|
||||||||
growing children: The University of Saskatchewan |
То же касается и воспалительных заболеваний кишечника |
|
||||||||
Bone Mineral Accrual Study. J Bone Miner Res. 1999; |
(ВЗК). В частности, высокая заболеваемость ВЗК и целиа- |
|
||||||||
14:1672–1679. |
|
|
|
|
|
|
кией наблюдается у людей, проживающих в северных широ- |
|
||
35. Bianchi M. L. Osteoporosis |
in children and |
adolescents. |
тах (например, в Великобритании и Скандинавии, на севере |
|
||||||
Bone. 2007;41(4):486–495. |
|
|
|
|
|
США), по сравнению с южанами, что связано с различиями |
|
|||
36. Мансурова Г. Ш., Мальцев С. В. Остеопороз у |
детей: |
в продолжительности и интенсивности воздействия сол- |
|
|||||||
роль кальция и витамина D в профилактике и терапии. |
нечного света [9–11]. Показано, что люди, живущие вблизи |
|
||||||||
Практическая медицина. 2017;5(106):55–59. |
|
|
|
экватора, имеют низкий риск развития ВЗК, однако при |
|
|||||
37. Czech-Kowalska J., Czekuc-Kryskiewicz E., |
Pludowski P., |
переезде в страны с умеренным климатом риск развития |
|
|||||||
Zaniuk K., Jaworski M., Łuba A., Grzybowska K., Piłat K., |
этих заболеваний у них увеличивается [12]. |
|
|
|||||||
Dobrzanska A. The Clinical and Biochemical |
Predictors |
Эпидемиологические исследования |
демонстрируют |
|
||||||
of Bone Mass in Preterm Infants. PLoS One. 2016;11(11): |
наличие сезонных колебаний заболеваемости инфекци- |
|
||||||||
e0165727. |
|
|
|
|
|
|
онной (респираторные инфекции, грипп, сепсис) [13–15] |
|
||
38. Consensus Development Conference: Diagnosis, prophylaxis |
и аллергической природы [16–18], а также неинфекци- |
|
||||||||
and treatment of osteoporosis. Am J Med. 1993;94:646–650. |
онными заболеваниями [19–21], в значительной степени |
|
||||||||
39. Шилин Д. Е. Эпидемиология переломов в детском воз- |
обусловленных уровнем инсоляции и связанным с ним |
|
||||||||
расте: обоснование фармакологической |
коррекции |
изменением обеспеченности витамином D. |
|
|
||||||
дефицита кальция и витамина D. Педиатрия. Журнал |
В крупном исследовании, включившем 415 пациентов |
|
||||||||
имени Г. Н. Сперанского. 2007;86(3):70–79. |
|
|
|
|
с рассеянным склерозом, проводилась оценка статуса вита- |
|
||||
40. Goulding A., |
Jones I. E., |
Taylor R. W., |
Manning P. J., |
мина D в зависимости от сезона. По результатам исследова- |
|
|||||
Williams S. M. More broken bones: a 4-year double cohort |
ния установлено, что средняя концентрация 25(OH)D сыво- |
|
||||||||
study of young girls with and without distal forearm |
ротки среди пациентов составила 24,8 нг/мл. Максимальная |
|
||||||||
fractures. J Bone Miner Res. 2000;15:2011–2018. |
|
|
концентрация кальцидиола (32,2 нг/мл) зарегистрирована |
|
||||||
41. Skaggs D. L., |
Loro M. L., |
Pitukcheewanont P., |
Tolo V., |
в июле-августе, в то время как минимальные значения |
|
|||||
Gilsanz V. Increased body weight and decreased radial |
(17,2 нг/мл) отмечены в зимний период (январь-февраль) |
|
||||||||
cross-sectional dimension in girls with forearm fractures. |
(p < 0,001). Самый низкий показатель распространенности |
|
||||||||
25 |
||||||||||
J Bone Miner Res. 2001;16:1337–1342. |
|
|
|
|
рецидивов наблюдался в сентябре-октябре (28%), наиболь- |
|||||
|
|
|
|
|
||||||
42. Щеплягина Л. А., Моисеева Т. Ю. Минерализация кост- |
шая их частота отмечена в марте-апреле (47%) (p < 0,005). |
|
||||||||
ной ткани у детей. Рос. пед. журн. 2003;3:16–22. |
|
|
Таким образом, авторы сделали вывод, что пику распро- |
|
||||||
43. Landin L. A. Epidemiology of children’s fractures. J Pediatr |
страненности рецидивов предшествовал период, в котором |
|
||||||||
Orthop B. 1997;6:79–83. |
|
|
|
|
|
концентрация 25(OH)D была минимальна [19]. |
|
|||
44. Rizzoli R., Bonjour J.-P., Ferrari S.L. Osteoporosis, genetics |
В исследовании среди пациентов с ВЗК (79 пациентов |
|
||||||||
and hormones. J Mol Endocrinol. 2001;26:79–94. |
|
|
с язвенным колитом и 141 — с болезнью Крона) также оце- |
|
||||||
|
|
|
|
|
|
|
нивалась взаимосвязь между обеспеченностью витамином D, |
|
||
|
|
|
|
|
|
|
сезоном и оценкой качества жизни пациентов по шкале |
|
||
4.2. Внекостные |
|
|
|
|
|
ВЗК (Inflammatory bowel disease questionnaire, IBDQ) [20]. |
|
|||
(некальциемические) проявления |
Средняя концентрация кальцидиола сыворотки также была |
|
||||||||
выше в летнее/осеннее время, чем в зимне-весенний период |
|
|||||||||
низкой обеспеченности организма |
|
|||||||||
(28,2 ± 0,9 против 23,8 ± 1,1 нг/мл соответственно; р = 0,002) |
|
|||||||||
витамином D |
|
|
|
|
|
(рис. 4.3, А). Эта разница в концентрации кальцидиола сыво- |
|
|||
|
|
|
|
|
|
|
ротки между сезонами наблюдалась как среди пациентов |
|
||
Для осуществления внекостных |
|
|
|
с болезнью Крона (27,8 ± 1,2 нг/мл летом и 23,2 ± 1,3 нг/мл |
|
|||||
|
|
|
зимой; р = 0,01), так и у пациентов с язвенным колитом |
|
||||||
(некальциемических) эффектов требуются большие |
|
|
||||||||
|
(29,0 ± 1,5 нг/мл летом и 24,9 ± 2,0 нг/мл зимой; р = 0,10). |
|
||||||||
дозы холекальциферола, чем для проявления костных |
|
|
||||||||
|
При этом в зимний/весенний периоды наблюдалась зна- |
|
||||||||
(кальциемических) функций |
|
|
|
|
|
|||||
|
|
|
|
чительная корреляция между низкой оценкой качества |
|
|||||
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
жизни и концентрацией сыворотки витамина D у пациен- |
|
||
4.2.1. Роль витамина D в регуляции |
|
|
тов с язвенным колитом (r = 0,35, р = 0,02) и незначитель- |
|
||||||
|
|
ная зависимость между данными показателями у пациентов |
|
|||||||
иммунной функции организма |
|
|
|
|
с болезнью Крона (r = 0,17, р = 0,06) (рис. 4.3, Б). |
|
||||
Взаимодействие витамина D с компонентами |
|
|
Наивысший балл по шкале IBDQ определяет лучшие |
|
||||||
|
|
показатели качества жизни. |
|
|
||||||
иммунной системы и его место в иммунорегуляции |
Аналогичная зависимость наблюдается и среди паци- |
|
||||||||
В последние годы отмечено увеличение количества |
ентов с системной красной волчанкой [21]. Так, по данным |
|
||||||||
исследований, посвященных иммуномодулирующей функ- |
F. Dall’Ara и соавт. [21], медианы кальцидиола сыворотки |
|
||||||||
ции витамина D. При этом доказано, что роль витамина D |
крови летом и зимой среди больных системной красной |
|
||||||||
не ограничивается лишь влиянием на звенья врожденного |
волчанкой составили 29,4 и 21,6 нг/мл |
соответственно |
|
|||||||
иммунитета, а также распространяется и на адаптивный |
(р = 0,005). |
|
|
|||||||
иммунитет [1–4]. |
|
|
|
|
|
|
Анализируя зависимость между частотой встречаемости |
|
||
Для ряда заболеваний характерна географическая неод- |
атопического дерматита и сезоном рождения, K. Mesquita |
|
||||||||
нородность, выражающаяся увеличением их распространен- |
и соавт. пришли к выводу, что более высокая распространен- |
|
||||||||
ности по мере удаления от экватора на север и на юг. В част- |
ность регистрируется у детей, рожденных осенью и зимой, |
|
||||||||
ности, самый высокий показатель распространенности |
по сравнению с родившимися весной и летом [16]. |
|
||||||||
сахарного диабета 1-го типа приходится на северные регионы |
Исследованиями показано, что дата рождения с апреля |
|
||||||||
(Финляндия) [5]. В Канаде, особенно в регионах с длинны- |
по август связана с более низким риском развития болезни |
|
||||||||
ми зимами, широко распространена неадекватная обеспечен- |
Крона в старшем возрасте [22], возможно, из-за увеличения |
|
||||||||
ность витамином D: в этих провинциях отмечается высокий |
внутриутробного или материнского витамина D. Кроме того, |
|
||||||||
уровень заболеваемости системными заболеваниями соеди- |
были отмечены выраженные сезонные различия в частоте |
|
||||||||
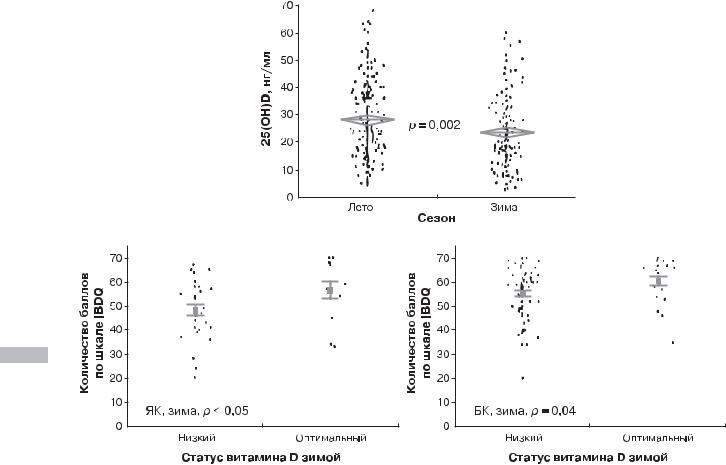
Рис. 4.3. Зависимость статуса витамина D от сезона (А) и оценка качества жизни в зависимости от статуса витамина D (Б) у пациентов с воспалительными заболеваниями кишечника [20]
А
26
Б
Примечание. ЯК — язвенный колит, БК — болезнь Крона.
первого эпизода и обострения ВЗК. Например, частота пер- |
активации происходит усиление синтеза фермента |
вой манифестации язвенного колита, по-видимому, достига- |
Cyp27B1, тем самым потенцируется превращение 25(ОН)D |
ет максимума в декабре, также осенью и зимой отмечались |
в 1,25(OH)2D [25, 26]. Концентрация 1,25(OH)2D, достиг- |
более высокие показатели рецидивов болезни Крона [12]. |
нутая за счет активации этого пути, необходима для синтеза |
|
важнейших факторов врожденного иммунитета — кате- |
Влияние витамина D на врожденный иммунитет |
лицидина и других антимикробных пептидов, с помощью |
На сегодняшний день рецепторы VDR обнаружены на |
которых усиливается антимикробная активность моноцитов |
поверхности практически всех иммунных клеток — CD4+ |
и макрофагов [27, 28]. Кроме того, 1,25(ОН)2D индуцирует |
и CD8+ лимфоцитах, В-лимфоцитах, нейтрофилах, анти- |
синтез IL1 непосредственно либо через усиление актив- |
генпрезентирующих клетках, в том числе макрофагах и ден- |
ности белков сигнального пути — с/EBP или ERK1/2 (+) |
дритных клетках [1, 2, 23], делая их восприимчивыми |
[29]. Поэтому для эффективного обезвреживания возбу- |
к витамин D-опосредованной модуляции (рис. 4.4). |
дителя посредством механизмов врожденного иммунитета |
Макрофаги представляют собой главный тип клеток |
в начале развития инфекционного процесса необходима |
системы мононуклеарных фагоцитов, их условно подразделя- |
оптимальная концентрация 1,25(OH)2D [23]. |
ют на две категории — М1- и М2-макрофаги. Макрофаги пер- |
Врожденные лимфоидные клетки. Этот термин отно- |
вого типа (М1) в ответ на действие антигенов вырабатывают |
сится к врожденной клеточной популяции с общим фено- |
провоспалительные медиаторы, такие как NO, TNF, IL23, IL12, |
типом и функциональной характеристикой. Врожденные |
IL1 , а также способствуют дифференцировке Т-хелперов |
лимфоидные клетки не имеют реаранжированного анти- |
(Th) до Тh1 и Th17 для участия в иммунном ответе. В свою |
генспецифического рецептора, и поэтому способны реагиро- |
очередь, макрофаги второго типа (М2) смещают баланс Th |
вать на широкий спектр сигналов. Они играют важную роль |
в направлении дифференцировки Th2, производят противо- |
в тканевом гомеостазе, формировании лимфоидной ткани |
воспалительный цитокин IL10, играющий важную роль в огра- |
и ее восстановлении [30]. |
ничении иммунных реакций, что крайне важно для заживле- |
Особенность «нетрадиционных» T-лимфоцитов ( T- |
ния ран и восстановления тканевого гомеостаза [23, 24]. |
лимфоциты, интраэпителиальные) заключается в экспрес- |
Макрофаги, являясь носителями VDR, демонстрируют |
сии гетеродимерного Т-клеточного рецептора, составленного |
высокую чувствительность к витамину D. Активные формы |
в отличие от традиционного -рецептора из - и -цепи [31]. |
витамина D способны влиять на макрофагальную диффе- |
На поздних стадиях инфекции действие витамина D |
ренцировку и активацию двумя путями. На ранних стади- |
в большей степени заключается в ограничении и регу- |
ях инфекции 1,25(ОН)2D стимулирует дифференциацию |
ляции интенсивности иммунного ответа [32]. Противо- |
моноцитов в макрофаги [25]. Кроме того, посредством toll- |
воспалительный эффект 1,25(ОН)2D характеризуется |
like рецепторов (TLR) или интерферон- -индуцированной |
снижением выработки макрофагами провоспалительных |
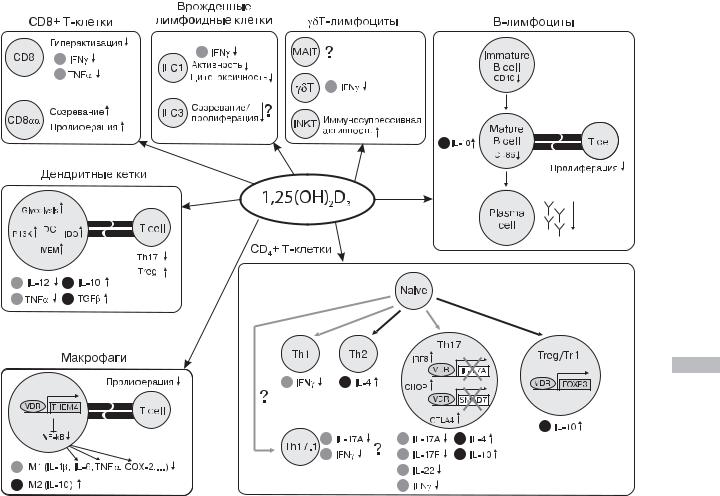
Рис. 4.4. Влияние активных форм витамина D на функционирование иммунной системы [23]
27
Примечание. Светлые точки представляют провоспалительные цитокины, темные точки — противовоспалительные цитокины. Светлые стрелки указывают на снижение дифференциации, темные стрелки — на усиление дифференциации.
факторов (IL1 , IL6, TNF, RANKL, NO) на фоне увеличения |
При этом происходит повышение синтеза противовоспа- |
продукции противовоспалительного IL10 [33, 34]. Эти изме- |
лительных IL10 и хемокина CCL22, ответственных за сти- |
нения, по-видимому, обусловлены стимуляцией дифферен- |
муляцию противоположных по эффектам субпопуляций |
цировки макрофагов M2 с ограничением распространения |
лимфоцитов Th2 и Treg [35, 36]. Следует отметить, что |
фенотипа М1, тем самым — восстановлением баланса между |
кальцитриол может вызывать этот толерогенный эффект |
этими подтипами клеток [23]. |
в отношении дендритных клеток только при обработке им |
Воздействие на макрофаги — важный, но не единствен- |
до их созревания, после созревания дендритных клеток спо- |
ный механизм влияния активных метаболитов витамина D |
собность 1,25(ОН)2D оказывать на них воздействие суще- |
на клетки иммунной системы: его концентрация также важ- |
ственно снижается [23, 36–38]. |
на и для других антигенпрезентирующих клеток. |
Дендритные клетки, как и макрофаги, экспрессируют |
Дендритные клетки представляют собой одно из наибо- |
на своей поверхности фермент CYP27B1, что позволяет |
лее важных звеньев в механизме иммунного ответа и регу- |
им генерировать высокую локальную концентрацию каль- |
ляции гомеостаза иммунной системы. Термином «ден- |
цитриола, которая необходима для модуляции иммунного |
дритные клетки» обозначаются несколько субпопуляций |
ответа [23, 32]. |
клеток иммунной системы с различными, а порой и карди- |
|
нально противоположными свойствами. Важнейшей функ- |
Роль витамина D в механизмах адаптивного |
цией дендритных клеток является презентация антигена |
иммунитета |
Т-лимфоцитам. Показано, что дендритные клетки в отли- |
Ранее было отмечено, что витамин D способен регули- |
чие от других антигенпрезентирующих клеток способны |
ровать реакции не только врожденного, но и приобретен- |
праймировать наивные Т-лимфоциты, вовлекая последние |
ного иммунного ответа. Однако исследователи отмечают |
в процесс первичного иммунного ответа [23]. |
трудности при определении его прямого воздействия на T- |
Дендритные клетки, дифференцированные in vitro |
и B-лимфоциты, так как большинство эффектов носят опос- |
из моноцитов или клеток костного мозга, в присутствии |
редованный характер, и проявляются через функциониро- |
1,25(ОН)2D способны оставаться в незрелом состоянии, что |
вание антигенпрезентирующих клеток. Несмотря на это, |
характеризуется снижением выработки провоспалительных |
было доказано, что количество VDR на CD4+ лимфоцитах |
факторов, таких как IL12, IL23 и TNF , тем самым снижая |
коррелирует со степенью их дифференцировки, что говорит |
последующую активацию Th1- и Th17-лимфоцитов, играю- |
о потенциальной восприимчивости Th к витамину D [23, 39]. |
щих важную роль в патогенезе аутоиммунных заболеваний. |
Наивные Т-лимфоциты содержат небольшое количество |
|
рецепторов, в то время как зрелые формы отличаются высо- |
CD4+ Т-клетки представляют собой гетерогенную группу, |
|
|
ким уровнем экспрессии VDR [1]. |
включающую Th1, Th2, Th17 и Treg клетки. При нормальном |
|
|
B-лимфоциты известны своей важнейшей ролью в |
иммунном ответе клетки Th1 важны для борьбы с внутри- |
|
|
иммунном ответе за счет продукции антител, они также |
клеточными патогенами, Th2 — с гельминтными инфек- |
|
|
способны презентировать антиген и секретировать цитоки- |
циями, Th17 — с внеклеточными патогенами и грибами; |
|
|
ны. Кроме этого, В-клеткам принадлежит решающая роль |
Treg опосредуют иммунологическую толерантность к ауто- |
|
|
в продукции аутоантител, что становится актуально при |
антигенам и безвредным чужеродным антигенам, таким как |
|
|
нарушениях иммунной регуляции и имеет определяющее |
пищевая и кишечная микробиота. Кроме того, они контро- |
|
|
значение в патогенезе аутоиммунных заболеваний [40]. |
лируют иммунный ответ через различные механизмы, вклю- |
|
|
Показано, что VDR связывается с промоторной обла- |
чая секрецию противовоспалительных медиаторов, таких |
|
|
стью генов, участвующих в работе иммунной системы в кле- |
как IL10 и трансформирующий фактор роста (TGF ) [44]. |
|
|
точной линии лимфобластных клеток, что свидетельству- |
Влияние витамина D на Т-клетки, опосредованное |
|
|
ет о восприимчивости В-лимфоцитов к витамину D [23]. |
кальцитриолом, заключается в регуляции их развития |
|
|
Продемонстрировано, что 1,25(OH)2D снижает пролиферацию |
и миграции. Прямыми объектами для активных форм вита- |
|
|
В-клеток, индуцирует их апоптоз. Это ингибирование диффе- |
мина D являются Th1 и Th2 клетки [23]. Но влияние вита- |
|
|
ренцировки может включать в себя предотвращение ядерной |
мина D на Th-лимфоциты оценивается неоднозначно. Так, |
|
|
транслокации NF-кB p65 (транскрипционный фактор) и бло- |
некоторые авторы отмечают ингибирование дифференци- |
|
|
кирование таким образом сигнального пути преобразования |
ровки Th0 в Th17 с одновременным уменьшением экспрес- |
|
|
костимулирующих молекул CD40 [40]. Кроме того, кальцитри- |
сии IL17 и IL21 в них, а также снижение дифференциа- |
|
|
ол индуцирует хемокиновый рецептор CCR10 на плазматиче- |
ции Th0 в Th1 с уменьшением продукции типичных для |
|
|
ских клетках, способствуя их миграции в очаги воспаления [41]. |
Th1 провоспалительных цитокинов (TNF , IFN , IL12) [10]. |
|
|
Имеется достаточно данных, доказывающих роль |
Добавление кальцитриола к CD4+ Т-клеткам способствует |
|
|
1,25(ОН)2D в снижении выработки антител [23, 42]. Инте- |
снижению экспрессии IL6 — кофактора, стимулирующего |
|
|
ресно, что наличие антинуклеарных антител коррелирует |
Th17 клетки, играющие важную роль в развитии аутоиммун- |
|
|
со снижением уровня сывороточного кальцидиола даже |
ных заболеваний [45]. В то же время другие исследователи |
|
|
у здоровых людей без системной красной волчанки. При |
демонстрируют отсутствие действия витамина D на Th1 [46]. |
|
|
этом сапплементация рациона препаратами холекальцифе- |
Стимулирование Th2 витамином D приводит к увеличе- |
|
28 |
|||
рола уменьшает титр этих аутоантител [43]. |
нию выброса противовоспалительных цитокинов IL4, IL5, |
||
|
|||
|
В целом, оказываемый кальцитриолом эффект на |
IL10 [10]. Ряд исследований демонстрирует, что воздей- |
|
|
В-клетки изучен не до конца. В настоящее время предпола- |
ствие витамина D приводит к увеличению пула Treg, а так- |
|
|
гается, что нормальный уровень 1,25(ОН)2D за счет продук- |
же способствует индукции синтеза IL10 [23, 46]. |
|
|
ции IL10 предотвращает их дифференцировку в плазмати- |
|
|
|
ческие клетки и тем самым снижает продукцию аутоантител |
Полиморфизм генов VDR как фактор риска |
|
|
[23]. Для подтверждения или опровержения этой гипотезы |
заболеваемости |
|
|
в настоящее время недостаточно данных, поэтому требуют- |
В настоящее время появляется все больше исследова- |
|
|
ся дальнейшие исследования. |
ний о полиморфизме генов VDR как о возможном факто- |
|
|
Исторически сложилось мнение, что дендритные клетки |
ре риска, обусловливающем предрасположенность к ряду |
|
|
были основной мишенью для витамина D, и что его эффек- |
инфекционных и неинфекционных заболеваний. |
|
|
ты, наблюдаемые в отношении Т-клеток, опосредовались |
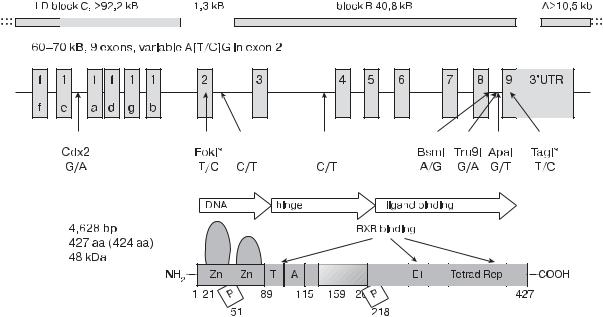
Ген VDR расположен на 12-й хромосоме (12q13.11). Он |
|
|
через дендритные клетки. Однако теперь стало ясно, что при |
имеет 11 экзонов и содержит четыре полиморфных области, |
|
|
воздействии витамина D различные популяции Т-клеток, |
три из которых расположены на конце гена (3’UTR) и пред- |
|
|
включая CD4+, CD8+ и TCR («нетрадиционные» Т-лим- |
ставлены рестрикционными фрагментами BsmI (rs1544410), |
|
|
фоциты), экспрессируют VDR, что делает Т-клетки прямой |
ApaI (rs 7975232) (интрон 8) и TaqI (rs 731236) (экзон 9) |
|
|
иммунологической мишенью для 1,25(OH)2D [44]. |
(рис. 4.5) [47]. Другая полиморфная область расположена |
Рис. 4.5. Структура гена рецептора витамина D [47]

в стартовом кодоне и представляет собой рестрикционный фрагмент FokI (rs 10735810) [47, 48].
Полиморфизмы рецептора витамина D изучались как потенциальные факторы, влияющие на развитие аутоиммунных заболеваний, включая рассеянный склероз [43, 49], аутоиммунный тиреоидит [50], ревматоидный артрит [51, 52], ВЗК [53], сахарный диабет 1-го типа [54, 55]; установлена роль полиморфизма генов VDR при бронхиальной астме [56, 57], а также инфекционных заболеваниях, включая туберкулез и ВИЧ-инфекцию [58–60].
Наиболее важные исследования о зависимости между развитием различных заболеваний и полиморфизмом генов рецептора витамина D представлены в табл. 4.7.
Очевидно, что не только витамин D и его активные метаболиты, но и экспрессия VDR в клетках органов-мишеней и клетках иммунной системы могут являться важными факторами, ассоциированными с рядом инфекционных и неинфекционных заболеваний.
4.2.2. Роль витамина D
в противоинфекционной защите
Помимо прямого модулирующего действия кальцитриола на различные клетки иммунной системы, имеется еще один очень важный механизм, обеспечивающий иммуно-
Таблица 4.7. Ассоциация генов, кодирующих VDR, с развитием ряда аутоиммунных и инфекционных заболеваний
Исследование |
Нозология |
Дизайн исследования |
Выявленный полиморфизм VDR |
|
|
|
|
|
|
|
|
Feng M. и соавт., 2013 [50] |
АИТ |
Метаанализ |
BsmI |
|
|
8 исследований |
TaqI |
|
|
||
|
|
|
|||
|
|
|
|
|
|
|
|
Метаанализ |
BsmI |
|
|
Zhang J. и соавт., 2012 [54] |
СД1 |
Доминантный вариант имеет выше риск |
|
|
|
57 исследований |
|
||||
|
|
СД1 на 30% (ОР 1,30; 95% ДИ 1,03–1,63) |
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
Метаанализ |
BsmI BB |
|
|
Sahin O. A. и соавт., 2017 [55] |
СД1 |
9 исследований |
BsmI Bb |
|
|
n = 1053 группа случая |
TaqI TT |
|
|
||
|
|
|
|||
|
|
n = 1017 группа контроля |
TaqI tt |
|
|
|
|
29 |
|||
|
|
|
|
|
|
|
|
|
Болезнь Крона: |
|
|
|
|
|
|
|
|
|
|
|
ApaI AA |
|
|
|
|
|
BsmI BB |
|
|
Wang L. и соавт., 2014 [53] |
ВЗК |
Метаанализ |
BsmI Bb |
|
|
19 исследований |
TaqI TT |
|
|
||
|
|
|
|||
|
|
|
TaqI Тt |
|
|
|
|
|
Язвенный колит: |
|
|
|
|
|
TaqI TT |
|
|
|
|
|
|
|
|
|
|
|
TaqI TT |
|
|
|
|
Метаанализ |
TaqI Тt |
|
|
|
|
BsmI BB |
|
|
|
|
|
12 исследований |
|
||
Tizaoui K. и соавт., 2015 [51] |
РА |
BsmI Bb |
|
|
|
n = 1703 группа случая |
|
||||
|
|
FokI FF |
|
|
|
|
|
n = 2635 группа контроля |
|
||
|
|
FokI Ff |
|
|
|
|
|
|
|
||
|
|
|
FokI ff |
|
|
|
|
|
|
|
|
|
|
Случай-контроль |
TaqI TT |
|
|
Di Spigna G. и соавт., 2016 [52] |
РА |
n = 40 группа случая |
|
||
TaqI tt |
|
|
|||
|
|
n = 40 группа контроля |
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Метаанализ |
TaqI TT |
|
|
|
|
11 исследований |
|
||
Huang J. и соавт., 2012 [43] |
РС |
TaqI Tt |
|
|
|
n = 2599 группа случая |
|
||||
|
|
TaqI tt |
|
|
|
|
|
n = 2816 группа контроля |
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Метаанализ |
ApaI Aa |
|
|
Tizaoui K. и соавт., 2015 [49] |
РС |
13 исследований |
ApaI aa |
|
|
n = 3300 группа случая |
ApaI AA — значительный фактор риска |
|
|
||
|
|
|
|||
|
|
n = 3194 группа контроля |
РС (p = 0,013) |
|
|
|
|
|
|
|
|
Zhao D. D. и соавт., 2017 [56] |
БА |
Метаанализ |
BsmI |
|
|
ApaI |
|
|
|||
|
|
|
|
||
|
|
|
|
|
|
|
|
Метаанализ |
ApaI |
|
|
|
|
9 исследований |
|
||
Han J. C. и соавт., 2016 [57] |
БА |
TaqI |
|
|
|
n = 2116 группа случая |
|
||||
|
|
FokI |
|
|
|
|
|
n = 1884 группа контроля |
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Метаанализ |
|
|
|
Cao Y. и соавт., 2016 [58] |
Туберкулез |
34 исследования |
|
|
|
n = 6525 группа случая |
FokI ff |
|
|
||
|
|
|
|||
|
|
n = 5669 группа контроля |
|
|
|
|
|
|
|
|
|
|
|
Случай-контроль |
|
|
|
Hu Q. и соавт., 2016 [59] |
Туберкулез |
n = 217 группа случая |
BsmI |
|
|
|
|
n = 338 группа контроля |
|
|
|
|
|
|
|
|
|
Nieto G. и соавт., 2004 [60] |
ВИЧ |
n = 185 группа случая |
FokI |
|
|
|
|
|
|
|
|
Примечание. АИТ — аутоиммунный тиреоидит, СД — сахарный диабет, ВЗК — воспалительные заболевания кишечника, РА — ревматоидный артрит, РС — рассеянный склероз, БА — бронхиальная астма, ВИЧ — вирус иммунодефицита человека.