Материал: Лекции Хомича Цитология
Лізосоми, які не увійшли в контакт з розщеплюваним матеріалом і їх ферменти перебувають у неактивному стані, називають первинними. Вторинні лізосоми формуються внаслідок злиття первинних лізосом з фагоцитозними й піноцитозними пухирцями (фаголізосоми) або зі зруйнованими частинами клітин (аутофагосоми). При цьому їхні ферменти активізуються і перетравлюють вміст пухирців. Третинні лізосоми (залишкові тільця) містять неперетравлені рештки біополімерів. Вміст цих лізосом з віком тварин може збільшуватись. Найбільше лізосом містять клітини, здатні до фагоцитозу. Руйнування білків відбувається не тільки у лізосомах, а й у поліпротеазних комплексах –
протеасомах.
Пероксисоми – субмікроскопічні органели загального призначення. Вони мають вигляд пухирців діаметром 0,2–0,5 мкм, які містять ферменти окиснення амінокислот, фермент каталазу і ферменти, що розщеплюють етиловий спирт, сечову кислоту та регулюють обмін ліпідів. Під час окиснення амінокислот утворюється пероксид Гідрогену, токсичний для клітин. Він руйнується ферментом каталазою. Найбільше пероксисом міститься в клітинах печінки, нирок та яєчників.
Немембранні органели. До них належать рибосоми, мікротрубочки, мікрофіламенти й клітинний центр (цитоцентр). Усі вони є органелами загального призначення.
Рибосоми – субмікроскопічні органели загального призначення, що утворюються в ядрі, а функціонують переважно в цитоплазмі. Їх вперше описав Д.Паладе. Вони мають вигляд округлих утворів діаметром 10–30 нм і складаються з двох субодиниць – великої й малої, які побудовані з білка та рибосомної РНК. Розрізняють поодинокі рибосоми і їх угруповання – полірибосоми. Вони можуть розміщуватись вільно в гіалоплазмі або приєднуватись до мембран ендоплазматичної сітки. Рибосоми беруть участь у синтезі білка. На них відбувається формування поліпептидів і білків. Вільні рибосоми синтезують білок, який використовується клітиною для власних потреб. Білок, синтезований рибосомами на мембранах ендоплазматичної сітки, потрапляє в порожнини її структур, транспортується в комплекс Гольджі і в складі секреторних гранул виводиться з клітини (секреторні білки) або використовується для формування лізосом і пероксисом.
33
Мікротрубочки – субмікроскопічні органели загального призначення. Вони мають вигляд довгих, прямих, нерозгалужених порожнистих циліндрів діаметром 24 нм, які в інтерфазнiй клітині формують сітку. Їхня стінка утворена щільно розміщеними 13 нитками субодиниць, які побудовані з молекул глобулярних білків тубулінів, здатних до полімеризації. Мікротрубочки дуже лабільні. У разі зміни умов навколишнього середовища (зниження температури) та під дією колхіцину їх формування припиняється і вони розпадаються на складові частини. Мікротрубочки формують цитоскелет, входять до складу органел (цитоцентр, війки, джгутики), беруть участь у внутрішньоклітинному транспорті мікропухирців, рибосом, мітохондрій та інших структур. Ними утворені базальні тільця. Під час репродукції клітин система мікротрубочок зникає і замість неї формується веретено поділу.
Мікрофіламенти – субмікроскопічні фібрилярні органели загального призначення. Вони мають товщину 5–17 нм і розміщені переважно в периферійній частині цитоплазми під плазмолемою та в цитоплазматичних виростах. Мiкрофiламенти побудовані зі скоротливих білків – актину, міозину, тропоміозину або альфа-актиніну і виконують опорно-рухову функцію.
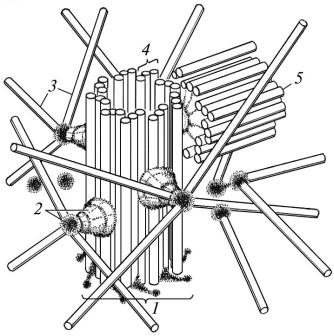
Клітинний центр (цитоцентр) – мікроскопічна органела за-
гального призначення. Її вперше описав В.Флемінг у 1875 році. Вона складається з двох центріолей (диплосома) і центросфери (рис. 6). Електронномікроскопічними дослідженнями встановлено, що центріолі мають вигляд порожнистих циліндрів діаметром 0,2 мкм і завдовжки 0,3–0,5 мкм, стінка яких утворена дев’ятьма триплетами паралельно розміщених мікротрубочок, які сполучаються між собою спеціальними структурами – “ручками”. Останні утворені білком динеїном, якому належить провідна роль у реалізації руху центріолей. Одну з центріолей називають материнською, другу – дочірньою. Материнська центріоля має додаткові структури – сателіти (місця відходження від неї мікротрубочок) і додаткові мікротрубочки. Ці мікротрубочки відходять від центріолі в радіальному напрямку і формують центросферу. В ділянці центросфери, гіалоплазма, крім мікротрубочок інших органел немає. Дочірня центріоля додаткових структур не має і розміщена під прямим кутом до материнської.
Центріолі є центрами формування мікротрубочок цитоплазми й мікротрубочок веретена поділу, яке забезпечує розходження хромосом під час поділу клітин. Вони ж формують базальні тільця аксонеми війок або джгутиків окремих різновидів клітин. Перед поділом клітини центріолі подвоюються.
34
|
Рис. 6. Цитоцентр (схема): |
|
1 |
— материнська центріоль; 2 — сателіт; 3 — мікротрубочки |
центросфери; |
4 |
— триплет мікротрубочок центріолі; 5 — дочірня центріоль |
|
|
О р г а н е л и с п е ц і а л ь н о г о п р и з н а ч е н н я . |
Д о н и х |
н а л е ж ать органели руху (війки, джгутики), мікроворсинки та мiкрофiбрили. Останні поділяють на тонофібрили, мiофiбрили i нейрофібрили.
Війки і джгутики – це тонкі вирости цитоплазми. Довжина війок становить 5–10 мкм, джгутиків – 150 мкм. Всередині війок і джгутиків є осьова нитка – аксонема, а в її основі – базальне тільце. Аксонема має вигляд циліндра, стінка якого утворена дев’ятьма парами мікротрубочок. У центрі циліндра знаходиться 10-та пара мікротрубочок. Усі мікротрубочки сполучені між собою, а також з базальним тільцем в єдину рухову систему. Мікроструктура базального тільця подібна до будови центріолі. Зміщення положення пар мікротрубочок аксонеми зумовлює рух війок і джгутиків. Війки характерні для епітеліальних клітин дихальних шляхів і маткової труби. Джгутик формує хвостовий відділ статевої клітини самця – сперматозоїда.
Мікроворсинки – це вирости цитоплазми, завдовжки 1 мкм, всередині яких знаходиться пучок мікрофіламентів. Вони збільшують поверхню клітини і добре розвинені в епітеліальних клітинах слизової оболонки кишок.
Тонофібрили, міофібрили і нейрофібрили – це різновиди мікрофібрил, які утворені мікротрубочками і мікрофіламентами та ха-
35
рактерні для клітин окремих тканин. Тонофібрили властиві епітеліальним клітинам. Вони формують їх цитоскелет. Міофібрили утворюють скоротливий апарат м’язових волокон і серцевих міоцитів. Нейрофібрили формують скелет нервових клітин.
Включення – непостійні компоненти цитоплазми. Вони накопичуються і зникають залежно від метаболічного стану клітин. Включення мають вигляд гранул, крапель і грудочок різного розміру. Їх поділяють на трофічні, секреторні, пігментні, вітамінні та екскреторні. До трофічних включень належать жири, вуглеводи й білки. Секреторні включення (секрет, інкрет) накопичуються в залозистих клітинах і є продуктами їх діяльності. Пігментні включення можуть бути екзогенного (каротин, барвники, часточки пилу) і ендогенного (гемоглобін, білірубін, меланін та ін.) походження. Наявність пігментів зумовлює колір клітин і тканин. Екскреторні включення – це продукти метаболізму, які шкідливі для клітин. Вони виводяться з клітин і організму. Вітамінних включень багато в клітинах печінки.
Ядро є складовою частиною клітин (рис.7). Разом з цитоплазмою вони утворюють єдину взаємодоповнювану систему, що перебуває в стані динамічної рівноваги. Клітина не може довго існувати і функціонувати без ядра – вона гине. Однак і ядро не здатне до самостійного існування без клітини.
Більшість клітин мають одне ядро, але бувають двоядерні (20% клітин печінки) і багатоядерні (мегакаріоцити, остеокласти) клітини. Форма ядер різноманітна і залежить здебільшого від форми клітин. Вони можуть бути округлими, паличкоподібними, кільцеподібними, овальними або сегментованими. Розмір ядер коливається в широких межах (3–40 мкм). Він залежить від особливостей клітин, їх віку та функціонального стану. Для окремих клітин крові (лімфоцити) характерне велике ядро, об’єм якого більший від об’єму цитоплазми. Ядра молодих клітин більші, ніж зрілих. Розмір ядер збільшується у клітин, що активно функціонують.
Ядра завжди розміщені в певних місцях клітин і бувають у двох станах – мітотичному та інтерфазному. Для кожного виду клітин властиве постійне співвідношення між об`ємом ядра і цитоплазми – ядер-
но-цитоплазматичне співвідношення (індекс Гертвіга).
Хімічний склад ядра подібний до такого цитоплазми. Однак у ядрі знаходиться майже вся ДНК, багато глобулярних білків і менше, ніж у цитоплазмі, РНК та ліпідів.
36
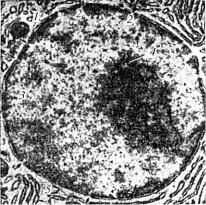
Рис. 7. Електронна мікрофотографія ядра кровотворної клітини ( 16000):
1 — оболонка ядра; 2 — пори в оболонці; 3 — грудочки хроматину; 4 — ядерце; 5 — гранулярна ендоплазматична сітка (за Фаусетом)
Ядро виконує функції, які можна об’єднати у дві групи. Перша група – це збереження спадкової (генетичної) інформації. До неї входять такі функції: підтримання сталої структури ДНК, редуплікація молекул ДНК, розподіл генетичного матеріалу під час поділу клітин та рекомбінація генетичного матеріалу в процесі мейозу. Друга група функцій реалізує генетичну інформацію – утворює апарат білкового синтезу. Це синтез усіх видів РНК і побудова рибосом. Таким чином, у ядрі міститься, функціонує і самовідтворюється генетичний матеріал.
Інтерфазне ядро складається з чотирьох компонентів: оболонки, нуклеоплазми, хроматину і ядерця.
Ядерна оболонка (нуклеолема) утворена двома клітинними мембранами завтовшки 7–8 нм, які відокремлені одна від одної перинуклеарним проміжком завширшки 20–60 нм. Зовнішня мембрана за своїми структурними особливостями подібна до мембрани гранулярної ендоплазматичної сітки. На ній також розміщені рибосоми, і вона може безпосередньо переходити в мембрану ендоплазматичної сітки. Внутрішня мембрана пов’язана з хроматином ядра. Через певні проміжки периметра ядра мембрани перериваються і переходять одна в одну. У цих місцях утворюються пори – отвори в нуклеолемі – діаметром 80–90 нм. Через пори відбувається транспорт речовин з ядра в цитоплазму і навпаки. Регуляція транспорту здійснюється комплексом пори, який утворений глобулярними й фібрилярними білками. Глобулярні білки утворюють гранули діаметром 25 нм. Вони розташовані по периметру стінки пори у три ряди. У кожному ряду є 8 гранул. У центрі пори міститься центральна гранула. Від гранул відходять фібрили, які сходяться у центрі пори і утворюють своєрідну діафрагму
37