Материал: Лекции Хомича Цитология
тактах плазмолема і цитоплазма однієї клітини утворюють пальцеподібні вирости, які заглиблюються у відповідні заглибини сусідньої клітини. Між плазмолемами контактуючих клітин зберігається щілина такої ж ширини як і у простому контакті.
Контакти за типом десмосом забезпечують максимальну міцність міжклітинних сполучень. Вони характерні для епітеліоцитів. У цьому контакті міжклітинна щілина завширшки 25–30 нм заповнена мікрофібрилами які проникають через плазмолеми у кортикальний шар цитоплазми сусідніх клітин і з`єднуються з пластинками прикріплення (скупчення фібрил). Напівдесмосомами з`єднуються епітеліоцити з базальною мембраною. Вони мають тільки одну пластинку прикріплення.
Щільний замикальний контакт переважно властивий епітеліоцитам, які вкривають слизову оболонку шлунка і кишечнику. Завдяки цьому контакту міжклітинні щілини ізолюються від зовнішнього середовища. У цьому контакті ділянки плазмолеми апікальних полюсів сусідніх епітеліоцитів максимально зближені. При цьому глікокалікс, білки і гідрофільні полюси молекул ліпідів плазмолеми контактуючих клітин утворюють єдиний шар.
Щілинний контакт або нексус забезпечує безпосередній транспорт молекул речовин між контактуючими клітинами. У ділянках таких контактів є міжклітинна щілина завширшки 2–4 нм, а в плазмолемі суміжних клітин особливі структури – конексони, які мають канали. У ділянці міжклітинної щілини конексони контактуючих клітин з`єднуються кінець у кінець. Внаслідок цього утворюються безпосередні зв`язки між клітинами.
Синапси – це контакти нервових клітин, а також нервових клітин з іншими структурами. Вони служать для односторонньої передачі нервового імпульсу. Будова синапсів, їх різновиди та особливості функціонування будуть описані при розгляданні нервової тканини.
Цитоплазма клітини складається з гіалоплазми, органел і вклю-
чень.
Г і а л о п л а з м а – це найбільш рідка частина цитоплазми. Вона становить близько 50% загального об’єму цитоплазми. До складу гіалоплазми входить вода з розчиненими в ній мінеральними та органічними речовинами (цитозоль). Високомолекулярні білкові речовини гіалоплазми у вигляді ниток утворюють ніжну суцільну тривимірну сітку, яка формує матрикс (строму) клітин. У гіалоплазмі є велика кількість ферментів, що беруть участь у процесах метаболізму, інформаційна і транспортна РНК. Через гіалоплазму здійснюється
28
внутрішньоклітинний транспорт речовин. У ній накопичуються запасні поживні речовини.
О р г а н е л и – частинки цитоплазми, які мають тільки їм властиву будову і спеціалізовані на виконанні певних функцій. Їх поділяють на мікроскопічні та субмікроскопічні, загального призначення й спеціальні, мембранні та немембранні. Мікроскопічні органели видимі під світловим мікроскопом, а субмікроскопічні можна побачити за допомогою електронного мікроскопа. Органели загального призначення є в усіх клітинах, а спеціального – в окремих різновидах клітин. Мембранні органели оточені клітинною мембраною, якої немає у немембранних органел.
М е м б р а н н і о р г а н е л и являють собою частини цитоплазми, відмежовані від гіалоплазми клітинними мембранами. Вони мають специфічний для них вміст, який за своїми властивостями відрізняється від гіалоплазми. До мембранних органел належать мітохондрії, ендоплазматична сітка, комплекс Гольджі, лізосоми й пероксисоми.
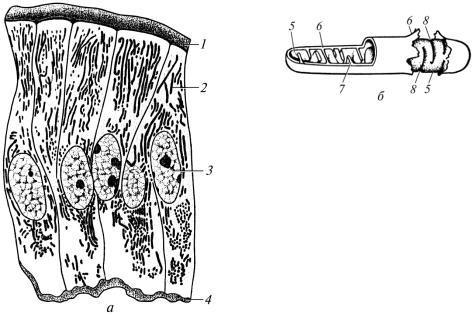
Мітохондрії – мікроскопічні органели загального призначення (рис. 4). Вони були відкриті в кінці ХІХ століття німецьким дослідником Ф.Альтманом. У них утворюється енергія, яка накопичується у молекулах АТФ і необхідна для життєдіяльності клітин. Мітохондрії беруть також участь у регуляції обміну води, газів, депонуванні іонів Са, Mg і P, продукуванні попередників стероїдних гормонів. Кількість мітохондрій у різних клітинах дуже варіабельна, від кількох до тисячі, що залежить від інтенсивності обмінних процесів.
|
|
Рис. 4. Мітохондрії: |
||
а — |
мітохондрії |
простого |
облямівкового |
|
епітелію |
кишки |
(світлова |
мікроскопія); |
|
б — |
схема |
будови |
мітохондрії (електронна |
|
мікроскопія); 1 — облямівка; 2 — мітохондрії;
3 |
— ядро; 4 — базальна мембрана; |
5 |
— внутрішня мембрана; 6 — зовнішня мем- |
брана; 7 — кристи; 8 — вигляд впинань (кристи) із зовнішньої поверхні внутрішньої мембрани
мітохондрій виявлено в клітинах печінки, нервових клітинах, міоци-
29
тах та м’язових волокнах. Мітохондрії функціонують близько 20 діб, після чого утилізуються лізосомами. Нові мітохондрії утворюються шляхом поділу або брунькуванням.
Під світловим мікроскопом мітохондрії мають вигляд нитко-, паличко- і зерноподібних структур завтовшки 0,2–2 мкм і завдовжки 1–10 мкм (рис. 4, а). Електронномікроскопічними дослідженнями встановлено, що мітохондрії обмежені двома мембранами завтовшки близько 7 нм (рис. 4, б). Між мембранами є проміжок завширшки 10– 20 нм. Зовнішня гладенька мембрана відділяє їх від гіалоплазми, а внутрішня обмежує вміст мітохондрій – мактрикс. Вона утворює численні складки, спрямовані всередину мітохондрій – кристи. Останні можуть мати вигляд пластинок або трубочок. Внаслідок цього мітохондрії ділять на пластинчасті та трубчасті. На внутрішній поверхні внутрішньої мембрани мітохондрій розміщені мітохондріальні субодиниці грибоподібної форми, в яких є ферменти, що забезпечують процес фосфорилювання. В матриксі мітохондрій містяться ферменти циклу Кребса та окиснення жирних кислот. У матрикс із гіалоплазми надходять продукти обміну білків, жирів, вуглеводів, Оксиген і АДФ. Продукти обміну білків, жирів і вуглеводів у результаті окиснення розщеплюються до води та вуглекислого газу, які виділяються з мітохондрій. При окисненні виділяється енергія, що забезпечує фосфорилювання (приєднання до АДФ залишку фосфорної кислоти). Внаслідок цього енергія накопичується в молекулах АТФ, які залишають мітохондрії. У місцях використання енергії від молекул АТФ відщеплюється залишок фосфорної кислоти. При цьому виділяється вільна енергія, яка забезпечує життєдіяльність клітин. У матриксі мітохондрій є також автономний апарат для синтезу мітохондріальних білків. Він включає власне мітохондріальні ДНК, РНК, рибосоми та ферменти, необхідні для їх синтезу і синтезу білка. Однак цей апарат через недостатню інформацію не може забезпечити синтез усіх структур мітохондрій, і синтез більшості ферментів забезпечує геном ядра клітини.
Ендоплазматична сітка – субмікроскопічна органела загального призначення, яку відкрив К.Портер у 1945 р. Вона складається із системи видовжених цистерн, трубочок, канальців і мішечків, які оточені клітинною мембраною та з`єднані між собою. Мембрана ендоплазматичної сітки може безпосередньо продовжуватись у зовнішню мембрану оболонки ядра і плазмолему. Ендоплазматична сітка є синтезуючою і транспортною системою клітини. У ній синтезуються різні органічні речовини, які накопичуються в порожнинах елементів сітки і транспортуються в різні ділянки клітин. В останніх, частини ендо-
30
плазматичної сітки з синтезованими речовинами відокремлюються і у вигляді пухирців надходять у гіалоплазму. Крім цього, ендоплазматична сітка ділить гіалоплазму на відсіки, у яких відбуваються специфічні біохімічні реакції.
Ендоплазматичну сітку поділяють на гранулярну і агранулярну. До зовнішньої поверхні мембран елементів гранулярної ендоплазматичної сітки прикріплюються у вигляді гранул рибосоми (рис. 5). У
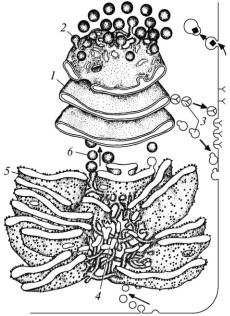
Рис. 5. Комплекс Гольджі та ендоплазматична сітка (схема):
1 — цистерни комплексу Гольджі; 2 — конденсуючі вакуолі; 3 — секреторні гранули; 4 — трубочки агранулярної ендоплазматичної сітки; 5 — цистерни гранулярної ендоплазматичної сітки; 6 — транспортні пухирці
зв’язку з цим функція цієї сітки полягає у синтезі білків як для потреб власне клітини, так і для виведення за її межі. Крім цього, у цій сітці відбувається синтез складових клітинної мембрани. Гранулярна ендоплазматична сітка дуже добре розвинена в клітинах, що синтезують білки. Ширина її складових може бути від 20 до 1000 нм. Зовнішня поверхня мембран елементів агранулярної ендоплазматичної сітки гладенька (рис. 5). У цій сітці відбувається синтез ліпідів і вуглеводів, детоксикація шкідливих продуктів обміну речовин, депонуванням іонів Са2+ (міоцити, м’язові волокна). Добре розвинена ця сітка в клітинах органів, які синтезують стероїдні гормони (надниркові залози, сім’яники). Її складові можуть мати ширину від 50 до 100 нм.
Комплекс Гольджі – мікроскопічна органела загального призначення, яка була вперше описана італійським гістологом К.Гольджі в 1898 році у нервових клітин. Під світловим мікроскопом, на препара-
31
тах оброблених азотнокислим сріблом, елементи комплексу Гольджі мають вигляд паличок і спірил. За допомогою електронної мікроскопії встановлено, що до його складу входить чотири компоненти: транспортні пухирці, вигнуті цистерни, які розміщені одна над одною (диктіосома), конденсуючі вакуолі й секреторні гранули (рис. 5). У комплексі Гольджі відбувається накопичення, конденсація і дозрівання продуктів, синтезованих в ендоплазматичній сітці, синтез поліцукридів, здійснюється зв’язок поліцукридів з білками. Тут також синтезується надмембранний комплекс оболонки клітини, утворюються лізосоми й пероксисоми. В окремих різновидах клітин може бути від однієї до кількох десятків цих органел. Найбільша кількість комплексів Гольджі є в секреторних клітинах.
Запитання до самоконтролю
1. Назвіть складові частини еукаріотної клітини. 2. Будова плазмолеми та її функції. 3. Як відбувається транспорт речовин у клітину та із неї? 4. Що зумовлює рецепторну функцію плазмолеми? 5. Що таке фагоцитоз і піноцитоз? 6. Які є різновиди екзоцитозу? 7. Види контактів. 8. Що таке гіалоплазма? 9. Які є критерії класифікації органел? 10. Будова і функції мітохондрій, ендоплазматичної сітки і комплексу Гольджі.
Лекція 4. Будова і функції еукаріотної клітини
Мікро- і субмікроструктура та функції лізосом, пероксисом, рибосом, клітинного центру, мікротрубочок і мікрофіламентів
.Органели спеціального призначення. Будова війок, джгутиків і мікроворсинок. Цитоплазматичні включення. Ядро та його функції. Складові ядра. Будова оболонки, нуклеоплазми, хроматину і ядерця ядра.
Лізосоми – субмікроскопічні органели загального призначення, які були відкриті у 1955 році Христіаном де Дювом. Вони мають вигляд пухирців діаметром 0,2–0,4 мкм, всередині яких міститься більше 60 гідролітичних ферментів – гідролаз. Завдяки цим ферментам лізосоми розщеплюють (перетравлюють) біополімери різного хімічного складу, через що їх називають органелами клітинного травлення. До таких біополімерів належать речовини, що надходять у клітину в результаті фагоцитозу й піноцитозу, а також зруйновані частини клітин.
32