Материал: Лекции Хомича Цитология
Колоїдні розчини протоплазми перебувають у двох фазах, які переходять одна в одну, – гель і золь. У фазі гелю колоїдний розчин щільніший. Його міцели в результаті агрегації формують просторові ґратки, в комірках яких знаходиться дисперсійне середовище. У фазі золю міцели відокремлюються одна від одної, колоїдний розчин стає текучим. Зміна фаз колоїдних розчинів протоплазми залежить від її функціонального стану та дії чинників зовнішнього середовища. Так, під час формування мітотичного веретена поділу клітин, утворення псевдоподій щільність колоїдів збільшується, а в разі зміни температури – зменшується.
Колоїдні системи протоплазми лабільні, їх білкові та ліпідні міцели формують структурну основу клітин. У разі ущільнення протоплазми утворюються нові білкові міцели, формуються зв’язки між ними, що лежать в основі регенеративних процесів клітин.
Якщо в колоїдний розчин потрапляють електроліти, настає процес коагуляції (зсідання). При цьому міцели злипаються і випадають в осад. Якщо дія коагулянту сильна, то процес коагуляції стає незворотним, що призводить до загибелі клітин. Явище коагуляції настає і тоді, коли міцели втрачають заряд.
У колоїдних розчинах може відбуватися процес коацервації. При цьому міцели втрачають зовнішній шар сольватної оболонки і з’єднуються за допомогою її внутрішніх шарів. У результаті цього утворюються великі агрегати – коацервати, які не сполучаються між собою. Коацервати мають рідку консистенцію. Їх формування часто спостерігається при потраплянні в клітини сторонніх речовин, що є захисною реакцією клітини.
Загальна характеристика еукаріотних клітин. Форма еу-
каріотних клітин різноманітна і визначається особливостями їх будови та функції, а також середовищем, що оточує клітину. Вона може бути кубічною, циліндричною, плоскою (епітеліоцити), кулястою (частина клітин крові), веретеноподібною (міоцити), пірамідною, зірчастою, з відростками (нервові клітини), багатокутною (клітини печінки) та ін. (рис. 2).
Клітини бувають рухливі й нерухомі. Нерухомі клітини мають сталу форму. Вони контактують між собою, утворюючи шари клітин. Клітини, здатні до активного руху, змінюють свою форму, наприклад нейтрофіли й фібробласти.
23
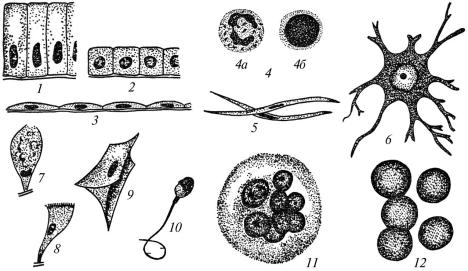
Рис. 2. Форма клітин:
1 — циліндрична; 2 — кубічна; 3 — плоска; 4 (а, б) — куляста; 5 — веретеноподібна; 6 — зірчаста; 7 — келихоподібна; 8 — війчаста; 9 — крилата;
Розміри клітин тварин не залежать від розмірів їх тіла. Найбільші клітини властиві амфібіям, у птахів і риб – клітини малих, а у ссавців – середніх розмірів. Поперечник (діаметр) клітини може бути
– від 4 до 150 мкм. Найбільші розміри мають статеві клітини самок – яйцеклітини та гігантські пірамідні клітини кори півкуль великого мозку, а найменші – клітини-зерна мозочка та малі лімфоцити.
Термін життя окремих видів клітин неоднаковий. Він триває доби, тижні (нейтрофіли), місяці (еритроцити), роки (клітини пам`яті). Окремі клітини (нейроцити) функціонують протягом всього життя індивідуума.
Клітини можуть бути одноядерні, двоядерні та багатоядерні. Кількість клітин в організмі ссавців величезна. Підраховано, що
загальна кількість клітин організму дорослої людини сягає 1014–1015. Еукаріотні клітини поділяють на соматичні й статеві. Будова
останніх з методичного погляду розглядається в розд. “Ембріологія”. Клітинні мембрани. Одним із головних компонентів структур-
ної організації клітин є клітинна мембрана, яку ще називають елементарною біологічною мембраною. За сучасними даними, вона побудована з подвійного шару молекул ліпідів, у який частково або повністю занурені молекули білків. Молекули ліпідів окремих шарів контактують між собою гідрофобними кінцями, а їх гідрофільні кінці спрямовані до периферії. Молекули білків можуть пронизувати всю товщу подвійного шару молекул ліпідів або значною мірою занурюватись у нього. Такі білки називають інтегральними. Частина молекул білків знаходиться на зовнішній поверхні ліпідів. Їх називають периферич-
24
ними, або адсорбованими, білками. Молекули білків залежно від функціонального стану структур клітин, обмежених мембранами, можуть переміщуватись у площині подвійного шару молекул ліпідів. Вони спеціалізовані у виконанні функцій. Залежно від цього молекули білків поділяють на структурні, ферментні, транспортні і рецепторні. Завдяки таким особливостям молекул білків клітинна мембрана не тільки відмежовує структури клітин від навколишнього середовища, а і бере участь у виконанні функцій цих структур.
Клітинні мембрани є внутрішньоклітинні і зовнішньоклітинні. До складу внутрішньоклітинних мембран належать мембрани, які утворюють оболонку ядра і мембрани, що оточують окремі органели. Зовнішньоклітинна мембрана одна – плазмолема. Клітинні мембрани можуть відрізнятись за хімічним складом.
Запитання для самоконтролю
1 .Що вивчає цитологія? 2. Хто і коли вперше побачив клітини? 3. Хто є творцем клітинної теорії? 4. Коли була створена клітинна теорія? 5. Основні положення клітинної теорії та їх обґрунтування. 6. Визначення клітини. 7. Які бувають клітини? 8. Як називають речовину, яка утворює клітину? 9. Хімічний склад протоплазми. 10. Фізико-хімічні властивості протоплазми. 11. Які форми можуть мати клітини і чим вони зумовлені? 12. Які розміри можуть мати клітини? 13. Чим утворена клітинна мембрана? 14. Функції клітинної мембрани. 15. Як ділять клітинні мембрани ?
Лекція 3. Будова і функції еукаріотної клітини
Частини еукаріотної клітини. Будова і функції плазмолеми. Цитоплазма. Склад цитоплазми. Гіалоплазма, органели і включення. Класифікація органел. Органели загального та спеціального призначення, мембрані та немембранні органели, мікро- і субмікроскопічні органели. Будова та функції ендоплазматичної сітки, мітохондрій і комплексу Гольджі.
Еукаріотна клітина складається з плазмолеми (оболонки), цитоплазми та ядра (рис. 3).
Плазмолема має товщину 10 нм. Її можна розглянути тільки за допомогою електронного мікроскопа. Вона обмежує клітину і склада-
25
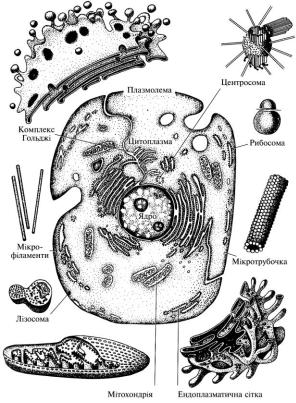
ється з трьох шарів: зовнішнього, середнього й внутрішнього. Зовнішній шар – глікокалікс або надмембранний комплекс, середній – клітинна мембрана і внутрішній – підмембранний комплекс. Будову клітинної мембрани описано вище. Глікокалікс утворений переважно вуглеводами – олігоцукридами, які утворюють з білками й ліпідами клітинної мембрани складні сполуки – відповідно глікопротеїди та гліколіпіди.
Рис. 3. Загальний план будови клітини (схема)
Вуглеводні кінці молекул цих сполук утворюють довгі розгалужені ланцюги. Підмембранний комплекс представлений мікрофіламентами й мікротрубочками, які розміщені в периферійній частині клітини і є частиною цитоскелета.
Плазмолема виконує численні функції. Основними з них є розмежувальна, транспортна, рецепторна, рухова, формування клітинних контактів та пристінного травлення. Розмежувальна і транспортна функції є взаємопротилежними і взаємодоповнюваними. Завдяки розмежуванню з навколишнім середовищем клітина зберігає свою індивідуальність, завдяки транспорту речовин здійснюються процеси обміну речовин як у самій клітині, так і між клітиною та середовищем, що її оточує. У результаті цього забезпечується сталість внутрішнього середовища клітини. Транспорт речовин всередину клітини нази-
26
вається ендоцитоз. Вода і розчинені в ній гази, окремі іони та дрібні молекули органічних речовин транспортуються в клітину i з неї шляхом дифузії (пасивний транспорт). Більшість іонів і невеликих молекул неорганічних та органічних речовин (солі, цукри, амінокислоти) транспортуються проти градієнта концентрації за участю особливих ферментів – пермеаз (активний транспорт супроводжується витрачанням енергії).Великі молекули органічних речовин та їх агрегати потрапляють у клітину в результаті фагоцитозу і піноцитозу. Фагоцитоз
– процес захоплення й поглинання клітиною великих часточок (бактерії, частинки пилу, фрагменти інших клітин), які мають щільну консистенцію. У процесі піноцитозу клітиною захоплюються й поглинаються макромолекулярні сполуки у вигляді розчинів. Часточки, що поглинаються, обволікаються плазмолемою і у вигляді мішечка втягуються всередину цитоплазми. Частини плазмолеми, які оточують поглинуті субстанції, втрачають зв’язок з плазмолемою клітини, внаслідок чого утворюються фагосоми й піноцитозні пухирці. Вони з’єднуються з лізосомами, під дією ферментів яких поглинуті часточки перетравлюються і засвоюються клітиною. Окремі поглинуті частинки можуть не з`єднуватись з лізосомами і виводиться з протилежної поверхні клітини. Таке явище називають трансцитоз.
Виведення клітиною речовин за межі цитоплазми називають екзоцитоз. Він має низку різновидів: секреція – виведення синтезованих продуктів; екскреція – виведення шкідливих або токсичних речовин; рекреція – виведення речовин, які не змінюють своєї хімічної структури в процесах метаболізму (вода, солі); клазматоз – видалення окремих структурних компонентів клітин. Механізми екзоцитозу прямо протилежні таким ендоцитозу.
Рецепторну функцію плазмолеми виконують глікокалікс і клітинна мембрана. Рух плазмолеми забезпечують елементи підмембранного комплексу, які здатні змінювати своє положення. Ферменти, що містяться в глікокаліксі плазмолеми ентероцитів, зумовлюють функцію пристінного травлення.
Плазмолема бере участь у формуванні міжклітинних контактів. При цьому вирішальну роль має глікокалікс. Контакти добре виражені в клітинах, які прилягають одна до одної (епітеліальні та ендотеліальні клітини). Залежно від особливостей будови, зумовлених їх функціями, міжклітинні контакти поділяють на такі види.
Прості контакти – між плазмолемами суміжних клітин знаходиться щілина завширшки 15–20 нм.
Зубчасті (інтердігітальні) контакти – збільшують площу контактуючих клітин і відповідно зміцнюють контакти. У таких кон-
27