Материал: Лекции Хомича Цитология
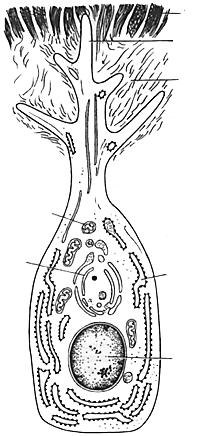
шоподібну форму і добре розвинені синтезуючі органели (рис. 60). Їх апікальна частина переходить у довгі відростки, які містяться у дентинових трубочках. Одонтобласти продукують колаген, з якого формуються колагенові волокна дентину і складові основної речовини. Через відростки одонтобластів у дентин надходять мінеральні і поживні речовини. На поверхню дентину починають відкладатися складові емалі, які синтезують амелобласти. У міру накопичення дентину і емалі амелобласти віддаляються від одонтобластів. У цьому разі пульпа і зовнішні клітини емалевого органа редукуються і після завершення утворення емалі вони разом з відростками амелобластів формують кутикулу емалі.
1
2
3
|
4 |
5 |
6 |
|
7
Рис. 60. Схема будови одонтобласта:
1 – дентин; 2 – відросток одонтобласта; 3 – предентин; 4 – мітохондрії; 5 – комплекс Гольджі; 6 – ендоплазматична сітка; 7 – ядро
Емаль відкладається тільки в ділянці коронки зуба. Амелобласти, які розташовані в ділянці майбутньої шийки зуба, незрілі. Вони розмножуються і заглиблюються в мезенхіму, утворюючи трубчасту епітеліальну кореневу піхву. Мезенхімоцити, які прилягають до її внутрішньої поверхні, диференціюються в одонтобласти, які продукують дентин кореня зуба. Таким чином дентин повністю оточує зу-
198
бний сосочок, з якого розвивається пульпа зуба. Із внутрішніх клітин зубного мішечка утворюється зубний цемент, із зовнішніх – періодонт. В останньому можуть бути клітинні елементи епітеліальної кореневої піхви, яка руйнується при диференціації зубного мішечка.
Посилення синтетичної активності клітин пульпи зуба супроводжується накопиченням у ній основної міжклітинної речовини, що тисне на сформовані тверді складові коронки зуба. Це призводить до її прорізування над поверхнею епітелію ясен.
Як відомо, частина зубів має молочних попередників. Молочні зуби розвиваються раніше постійних. Закладка постійних зубів відбувається з епітелію зубної пластинки позаду закладки молочних зубів. Спочатку молочні та постійні зуби містяться у спільній альвеолі. Дещо пізніше між ними виникає кісткова перетинка. Під час зміни зубів остеобласти руйнують її та корінь молочного зуба. Останній видаляється з альвеоли. Під дією тиску, який виникає у пульпі постійного зуба, внаслідок активації синтетичної діяльності її фібробластів, його коронка виштовхується над поверхнею ясен.
Слинні залози належать до травних залоз. Вони продукують секрет – слину, яка на 90 % утворена водою, а також мінеральними та органічними речовинами. До мінеральних речовин слини належать переважно іони натрію, кальцію та калію; до органічних речовин – бі- лки-ферменти і слиз. У слині виявляються лейкоцити, злущені епітеліоцити і екскреторні речовини (йод, креатин, сечова кислота тощо).
Слина зволожує корм, сприяє його механічній обробці, формуванню кормової грудки та її ковтанню. Ферменти слини (амілаза, мальтоза, гіалуронідаза, трипсинподібні тощо) беруть участь у хімічній обробці корму. Слина, завдяки наявності в ній лізоциму і лейкоцитів, виявляє бактерицидну дію. Зі слиною виділяються окремі проміжні і кінцеві продукти обміну речовин (залізо, йод, сечова кислота, креатин). У великих слинних залозах (застінних) утворюються біологічно активні речовини (гормони), які потрапляють у кров і впливають на обмін вуглеводів, мінеральних речовин, ріст епітелію тощо.
Слинні залози – це екзокринні залози. Тобто вони утворені секреторними відділами і протоками. Їх поділяють на пристінні та застінні. Перші розташовані у слизовій оболонці більшості органів, які формують стінки порожнини рота (губи, щоки, язик, м’яке піднебіння), а другі – за межами її стінок. Протоки усіх слинних залоз відкриваються у порожнину рота.
Застінних слинних залоз є три пари: привушна, піднижньощелепна і під’язикова. Їх секреторні відділи і протоки розвиваються із ектодерми, а сполучнотканинна строма – із мезенхіми. Сполучнотка-
199
нинна строма утворена пухкою волокнистою сполучною тканиною. Вона представлена капсулою, яка окутує залози зовні і перегородками, що відходять від неї. Перегородки поділяють залози на часточки. У сполучнотканинній стромі містяться кровоносні та лімфатичні судини, нервові вузли, волокна і закінчення, а в перегородках ще й міжчасткові вивідні протоки. У часточках розташовані секреторні відділи і внутрішньочасточкові вивідні протоки: вставні та посмуговані. Навколо складових часточок містяться ніжні прошарки пухкої волокнистої сполучної тканини з численними кровоносними судинами. Секреторні відділи застінних слинних залоз бувають трьох типів: серозні,
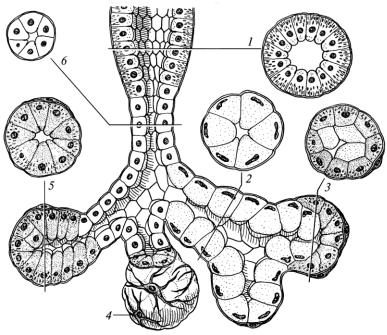
слизові та змішані (рис. 61).
Рис. 61. Схема будови секреторних відділів і внутрішньочасточкових проток
слинних залоз:
1 – посмугована протока; 2 – слизовий секреторний відділ; 3 – змішаний секреторний відділ; 4 – міоепітеліоцити; 5 – серозний секреторний відділ; 6 – вставна протока
Клітини серозних (білкових) секреторних відділів (сероцити) синтезують переважно білки-ферменти і глікопротеїн, який зв’язує та транспортує у слину імуноглобулін А, що утворюється у плазмоцитах волокнистої сполучної тканини, яка оточує секреторні відділи. У клітинах слизових секреторних відділів (мукоцитах) синтезуються слизоподібні білки і протеоглікани. До складу змішаних секреторних відділів належать сероцити і мукоцити.
Клітини секреторних відділів слинних залоз виділяють секрет за мерокриновим типом.
200
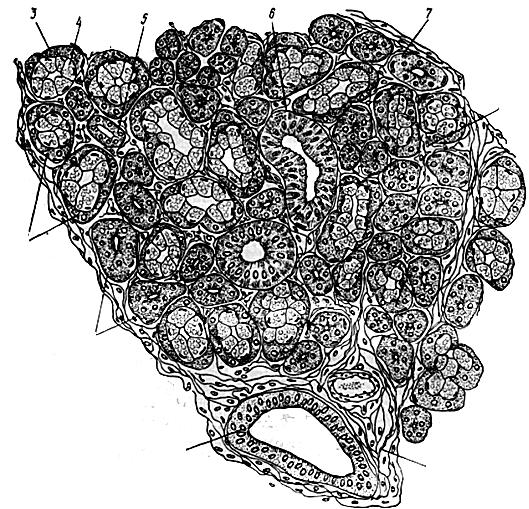
Піднижньощелепна залоза – складна, альвеолярно-трубчаста, розгалужена, з серозно-слизовим типом секрету. В її часточках є секреторні відділи серозного і змішаного типів (рис. 62).
3 |
|
|
|
|
|
6 |
|
7 |
|
4 |
|
5 |
|
|
|||
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
8
2
1
10
9
Рис. 62. Схема мікробудови піднижньощелепної слинної залози:
1 – серозний секреторний відділ; 2 – змішаний секреторний відділ; 3 – серозний півмісяць; 4 – слизові клітини змішаного секреторного відділу; 5 – вставна протока; 6 – посмугована протока; 7 – міоепітеліоцит; 8 – внутрішньочасточкова сполучна тканина; 9 – міжчасточкова сполучна тканина; 10 – міжчасточкова протока
Серозних відділів найбільше. Вони утворені сероцитами і міоепітеліоцитами, які оточені базальною мембраною. Сероцити мають конічну форму і базофільну цитоплазму. Ядро їх кулясте і розташоване в центрі клітини. У цитоплазмі містяться добре розвинені гранулярна ендоплазматична сітка і комплекс Гольджі. Просвіт серозних відділів дуже малий. Він продовжується у міжклітинні канальці, які знаходяться між сероцитами. Таким чином секрет із сероцитів виділяєть-
201
ся не тільки через їх апікальний полюс, а й через бічні поверхні. Міоепітеліоцити прилягають до сероцитів з боку їх основ. Це відросчасті клітини, у цитоплазмі яких є скоротливі структури. Скорочення міоепітеліоцитів сприяє виведенню секрету із секреторних відділів у протоки.
Змішані секреторні відділи утворені мукоцитами, сероцитами та міоепітеліоцитами, що оточені базальною мембраною. Мукоцити розташовані ближче до просвіту секреторного відділу. Вони крупні, мають конічну форму з широкою основою. Їх цитоплазма світла, містить багато гранул слизового секрету. Ядро мукоцитів при накопиченні секрету зміщується у базальну частину клітини і стає плоским. У цитоплазмі цих клітин добре розвинені агранулярна і гранулярна ендоплазматична сітка, комплекс Гольджі. Сероцити прилягають до основ мукоцитів і формують так звані білкові півмісяці. Ці клітини відрізняються від мукоцитів формою, центральним розміщенням ядра і базофільною цитоплазмою. Вони синтезують лізоцим, який, як відмічено вище, виявляє бактерицидну дію. Міоепітеліоцити прилягають до основ сероцитів.
Система проток піднижньощелепної залози починається вставними протоками. Вони є продовженням секреторних відділів. Їх стінка утворена шаром плоских або кубічних епітеліоцитів, до яких прилягають веретеноподібні міоепітеліоцити, оточені базальною мембраною. В епітеліоцитах вставних проток можуть бути білкові секреторні гранули. Із збільшенням віку тварин частина епітеліоцитів цих проток перетворюється на мукоцити. Міоепітеліоцити стінки вставних проток сприяють проштовхуванню секрету в посмуговані протоки. Посмуговані протоки формуються внаслідок злиття декількох вставних проток. Вони мають більший діаметр ніж вставні протоки. Їх стінка утворена шаром високих призматичних епітеліоцитів, які розташовані на базальній мембрані. Серед епітеліоцитів трапляються келихоподібні клітини і ендокриноцити. Для епітеліоцитів властива ацидофільна цитоплазма і базальна посмугованість. Остання зумовлена наявністю глибоких впинань плазмолеми базального полюса епітеліоцитів у їх цитоплазму. Між впинаннями розташовані численні мітохондрії. Наявність базальної посмугованості в епітеліоцитах посмугованих проток свідчить про їх участь у транспорті води і розчинів мінеральних речовин між вмістимим проток та міжклітинною речовиною волокнистої сполучної тканини, яка оточує їх. Посмуговані протоки дають початок міжчасточковим протокам. Їх стінка утворена двошаровим призматичним епітелієм, який знаходиться на базальній мембрані. Міжчасточкові протоки зливаються і утворюють головну протоку, яка утво-
202