Материал: Лекции Хомича Цитология
ються, формуючи нервову трубку, в центрі якої є канал. Біля бічних поверхонь нервової трубки розташовані нервові гребені, а в ділянці її головного кінця – плакоди. Стадію утворення нервової трубки нази-
вають нейруляцією.
Паралельно з нейруляцією відбувається диференціація ентодерми. На її дорсальній поверхні виділяється хордо-мезодермальний зачаток, який ділиться на хорду і мезодерму. Спочатку хорда має вигляд пластинки, краї якої в подальшому змикаються і вона перетворюється на суцільний клітинний тяж, розміщений під нервовою трубкою. Мезодерма міститься по боках хорди і представлена двома мішкоподібними випинаннями дорсальної стінки ентодерми, які з часом відділяються від неї. Після виділення зі складу ентодерми хорди та мезодерми її називають кишковою ентодермою (вторинною), яка формує кишкову трубку.
Диференціація зародкових листків i осьових органів відбувається, як описано вище.
Ембріогенез риб. Яйцеклітину риб називають ікринкою. Для більшості риб властиве зовнішнє запліднення. Представники окремих видів класу риб різняться за індивідуальним розвитком. Одні з них у процесі розвитку проходять стадію личинки, інші її не мають. Ці особливості розвитку пов’язані з особливостями їх ембріогенезу.
Риби, розвиток яких відбувається зі стадією личинки (дводишні кистепері, осетрові), мають мезо- і телолецитальні яйцеклітини. Такі яйцеклітини властиві й амфібіям. Відповідно ембріогенез цих риб такий самий, як і у амфібій.
Для риб, що розвиваються без стадії личинки (вищі костисті риби, акули), характерні полілецитальні яйцеклітини, як і для птахів. Ембріогенез цих риб подібний до ембріогенезу птахів. Однак для розуміння еволюції ембріогенезу потрібно знати, що вперше в ембріогенезі представників цих риб формуються позазародкові органи (жовтковий мішок).
Ембріогенез амфібій. Яйцеклітину амфібій, як і риб, називають
ікринкою (рис. 19).
Це мезо- і телолецитальна яйцеклітина, в ділянці анімального полюса якої знаходиться пігмент чорно-бурого кольору. Він поглинає теплові промені, що прискорює розвиток зародка.
Дроблення зиготи амфібій повне нерівномірне. Перші дві борозни дроблення проходять у меридіональній площині, як у ланцетника. Третя борозна дроблення проходить у широтній площині ближче до анімального полюса. В результаті зародок на цій стадії дроблення складається з чотирьох анімальних малих бластомерів (мікромерів) і
68
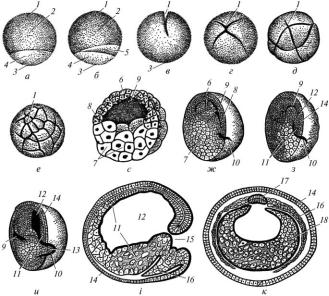
чотирьох вегетативних великих бластомерів (макромерів). Після цього дроблення відбувається почергово в меридіональній і широтних площинах. Однак дроблення бластомерів вегетативного полюса відстає від дроблення бластомерів анімального полюса, що призводить до порушення збільшення кількості бластомерів у геометричній прогресії. Крім того, дроблення відбувається також у тангенційній площині.
Рис. 19. Ембріогенез жаби (схема):
а — рання зигота; б — зигота через 2 год після запліднення; в, г, д, е — послідовні стадії дроблення; є — розріз бластули; ж, з, и, і (поздовжній розріз), к (поперечний розріз) — гаструляція; 1 — анімальний полюс; 2 — пігментована частина зиготи; 3 — вегетативний полюс; 4 — непігментована частина зиготи; 5 — сірий півмісяць; 6 — дах бластули; 7 — дно бластули; 8 — крайова зона; 9 — бластоцель; 10 — серпоподібна борозна; 11 — ентодерма; 12 — гастроцель; 13 — бластомери дна бластули; 14 — ектодерма; 15 — бластопор; 16 — зачаток мезодерми; 17 — зачаток хорди; 18 — кишкова ентодерма
У результаті дроблення утворюється амфібластула, бластодерма якої багатошарова i має нерівномірну товщину. У ділянці дна вона утворена макромерами, у ділянці даху – мікромерами. Порожнина амфібластули зміщена в бік даху.
Гаструляція починається інвагінацією і змінюється епіболією. В ділянці крайової зони поблизу дна бластодерма починає впинатися в порожнину амфібластули, внаслідок чого утворюється серпоподібна борозна. Однак цей шлях гаструляції в подальшому стає неможливим, оскільки переобтяжені жовтком макромери бластодерми дна амфібластули не можуть впинатися. У зв’язку з цим далі гаструляція відбувається за типом епіболії. Мiкромери даху та крайової зони амфібластули активно діляться і обростають макромери її дна. Внаслідок цьо-
69
го зародок стає двошаровим і складається з екто- і ентодерми. Ектодерма формується з бластомерів даху амфібластули, ентодерма – з бластомерів дна, крайової зони і частково даху.
Серпоподібна борозна подовжується по колу, її кінці зливаються і утворюється бластопор, який розміщений на задньому кінці зародка. В центрі бластопора знаходиться жовткова пробка, утворена бластомерами дна амфібластули. Формування бластопора відбувається внаслідок переміщення клітин презумптивних хорди й мезодерми. Клітини презумптивної хорди переміщуються через дорсальну губу бластопора. Вони прямують у головному напрямку і входять до складу ентодерми. Клітини презумптивної мезодерми переміщуються через вентральну і латеральні губи бластопора. Вони не з’єднуються з ентодермою і розміщені між нею та ектодермою.
У процесі ембріогенезу зачаток хорди виходить зі складу ентодерми, внаслідок чого формуюча кишкова трубка з дорсальної поверхні стає незамкненою і має вигляд жолоба. Завдяки активному розмноженню клітин крайових зон краї жолоба змикаються і формується кишкова трубка. Вентральна й латеральні ділянки кишкової трубки багатошарові. Шари клітин, розміщених поблизу порожнини кишки, утворюють жовткову ентодерму, яка використовується для живлення зародка, а найглибший – кишкову ентодерму, яка формує власне кишкову трубку.
Отже, у амфібій зачаток мезодерми не виділяється з ентодерми, як у ланцетника.
Переміщення клітинного матеріалу призводить до звуження бластопора, його жовткова пробка зникає у зв’язку із заглибленням бластомерів дна амфібластули всередину гаструли.
Утворення нервової трубки відбувається, як i у ланцетника, а диференціація зародкових листків та осьових органів – як описано вище.
Ембріогенез птахів. Яйцеклітина і, відповідно, зигота птахів полі- і телолецитальна. Запліднення птахів внутрішнє і відбувається в яйцепроводі. Його особливістю є поліспермія – проникнення в яйцеклітину до 300 сперматозоїдів. Однак з пронуклеусом яйцеклітини зливається пронуклеус тільки одного сперматозоїда. Явище поліспермії пов’язане з тим, що кортикальна реакція під час запліднення відбувається дуже повільно, внаслідок чого формування оболонки запліднення сповільнюється.
Дроблення зиготи птахів часткове. Перші дві борозни дроблення проходять у меридіональній площині перпендикулярно одна до одної. Потім дроблення відбувається почергово в меридіональній та широтних площинах. У результаті дроблення утворюється дискобласту-
70
ла. Її дах i крайова зона утворені зародковим диском, а дно – жовтком. Порожнина дискобластули щілиноподібна.
Дроблення й початок гаструляції відбуваються в яйцепроводі, де яйце перебуває впродовж 4–27 год. і вкривається третинною оболонкою. Отже, знесене яйце є зародком на ранньому етапі ембріонального розвитку. У знесеному яйці у зв’язку зі зміною температурних умов ембріогенез припиняється і поновлюється лише з початком насиджування або інкубації.
Гаструляція відбувається за типом деламінації та імміграції. В результаті цих процесів зародковий диск у знесеному яйці складається з внутрішнього та зовнішнього зародкових листків. У перші 12 год інкубації зародковий диск розростається на поверхні жовтка. Його центральна частина утворює зародковий щиток, з якого розвивається зародок, а периферична – позазародкову частину, з якої формуються позазародкові органи (плодові оболонки). Клітини зародкового щитка і частково позазародкової частини розміщуються над підзародковою порожниною, яка утворилася внаслідок використання ними частини жовтка та формують світле поле зародкового диска. Світле поле оточене темним полем, яке утворене клітинами позазародкової частини зародкового диска, що лежать безпосередньо на жовтку.
Узародковому диску здійснюється переміщення клітинного матеріалу презумптивної хорди й мезодерми. Воно відбувається двома потоками в напрямку до заднього кінця зародкового щитка, внаслідок чого він подовжується. У ділянці заднього кінця потоки клітинного матеріалу зливаються і прямують по серединній лінії зародкового щитка в передньому (головному) напрямку, формуючи клітинний тяж – первинну смужку. Вона закінчується потовщенням – первинним (гензенівським) вузликом. Через первинні смужку і вузлик проходить заглиблення (підвертання) клітинного матеріалу презумптивної хорди і мезодерми під ектодерму. При цьому в первинній смужці утворюється
первинна борозна, а в первинному вузлику – первинна ямка. Первинна ямка відповідає дорсальній губі бластопора, а первинна борозна – іншим губам бластопора. Через первинну ямку переміщується клітинний матеріал хорди. Він прямує в напрямку головного кінця зародка і називається головним відростком. Через первинну борозну переміщується клітинний матеріал мезодерми. У міру переміщення клітинного матеріалу первинна смужка вкорочується, і первинний вузлик зміщується до заднього кінця зародка.
Уході гаструляції зародкова частина зародкового диска відмежовується від позазародкової частини за допомогою тулубової складки. Вона формується внаслідок інтенсивного розростання клітин
71
ділянок усіх зародкових листків, що знаходяться на межі зародкової та позазародкової частин зародкового диска. Верхівка складки спрямована під зародок, внаслідок чого він підіймається над жовтком. При цьому краї зародкової ентодерми змикаються і формується кишкова трубка, яка сполучена з жовтком вузькою жовтковою протокою. Формування нервової трубки і диференціація зародкових листків та осьових органів відбувається так само, як у ланцетника та амфібій.
Запитання для самоконтролю
1. Що розвивається з зародкових листків і осьових органів? 2. Склад мезодерми та її диференціація. 3. Ембріогенез ланцетника. 4. Ембріогенез риб. 5. Ембріогенез амфібій. 6. Ембріогенез птахів.
Лекція 9. Ембріогенез птахів і ссавців
Позазародкові органи птахів. Стадії розвитку курчати. Ембріогенез плацентарних ссавців. Позазародкові органи ссавців. Періоди внутрішньоутробного розвитку ссавців.
Позазародкова частина зародкового диска складається з позазародкових екто-, енто- і мезодерми. Остання представлена тільки несегментованою частиною (спланхнотом).
Із позазародкової частини зародкового диска розвиваються тимчасові позазародкові органи – плодові оболонки. До них належать жовтковий мішок, амніон, сероза та алантоїс.
Першим з позазародкових органів розвивається жовтковий мішок. Він формується в результаті обростання жовтка позазародковими ентодермою та вісцеральним листком мезодерми. Поблизу зародка вони формують жовткову протоку, яка входить до складу пупка, і, як було зазначено вище, сполучає кишкову трубку із вмістом жовткового мішка. Процес обростання жовтка не відбувається до кінця, внаслідок чого жовтковий мішок знизу залишається відкритим. У цій ділянці жовток безпосередньо прилягає до білка. Жовтковий мішок виконує трофічну функцію, є органом ембріонального кровотворення та органом, у якому утворюються первинні статеві клітини – гоноцити. Ентодерма стінки жовткового мішка бере участь у ферментативному розщепленні жовтка та всмоктуванні поживних речовин у кровоносні судини мезодермального шару, які продовжуються в судини
72