Материал: Фарм производство БАР
Существует мнение, что природные интерфероны, которые являются смесью разных подклассов и видов, имеют более высокую терапевтическую эффективность в сравнении с рекомбинантными интерферонами. Так, например, природный α-интерферон может использоваться для лечения кондилом одинаковой с рекомбинантным интерфероном эффективностью в дозах в 4 раза меньших по сравнению с рекомбинантным интерфероном
Получение рекомбинантного интерферона. В настоящее время большинство препаратов интерферона получено методами генной инженерии. Применение рекомбинантного интерферона позволило расширить арсенал лекарственных средств для лечения очень многих тяжелых заболеваний: новообразований лимфатической системы и системы кроветворения (хронический миелолейкоз, кожная Т-клеточная лимфома, неходжкинская лимфома низкой степени злокачественности и др.); солидные опухоли (саркома Капоши у больных СПИДом, метастазирующая меланома, меланома после хирургической резекции и др.); вирусные заболевания (хронический активный гепатит В, гепатит С и др.).
Первым этапом создания рекомбинантного интерферона является выделение кДНК интерферонов. Для выделения генов или кДНК белков человека используют разные подходы. В ряде случаев выделяют нужный белок и определяют аминокислотную последовательность соответствующего участка молекулы. Исходя из этого находят кодирующую его нуклеотидную последовательность, синтезируют соответствующий олигонуклеотид и используют его в качестве гибридизационного зонда для выделения нужного гена или кДНК из геномных или кДНК-библиотек. Другой подход состоит в выработке антител к очищенному белку и использовании их для скрининга библиотек, в которых происходит экспрессия определенных генов. Для белков человека, синтезируемых преимущественно в какой-то одной ткани, кДНК-библиотека, полученная на основе мРНК, выделенной из этой ткани, будет обогащена последовательностью ДНК-мишени. Например, основным белком, синтезируемым клетками островков Лангерганса поджелудочной железы, является инсулин, и 70 % мРНК, выделенных из этих клеток, кодируют, именно его.
238
Однако принцип обогащения кДНК неприменим для тех белков человека, количество которых очень мало или место синтеза которых неизвестно. В этом случае понадобиться использовать другие экспериментальные подходы. Интерфероны (α, β, γ) – это природные белки, каждый из которых может найти свое терапевтическое применение. При выделении их кДНК пришлось разработать новый подход, позволяющий преодолеть трудности, связанные с недостаточным содержанием соответствующих мРНК и белков.
Процедура выделения кДНК интерферонов состояла в следующем:
1.Из лейкоцитов человека выделили мРНК и фракционировали её по размерам; провели обратную транскрипцию и встроили в сайт PstI плазмиды pBR322.
2.Полученным продуктом трансформировали Escherichia coli. Образовавшиеся 6000 клонов подразделили на 12 групп: по 512 клонов в каждой. Тестирование проводили на группе клонов, что позволило ускорить процесс их идентификации;
3.Каждую группу клонов гибридизовали с неочищенным препаратом интерферон-мРНК;
4.Из образовавшихся гибридов, содержащих клонированную ДНК и мРНК, выделили мРНК и провели её трансляцию в бесклеточной системе синтеза белка;
5.Определили интерферонную противовирусную активность каждой смеси, полученной в результате трансляции. Группы, проявившие интерферонную активность, содержали клон с кДНК, гибридизовавшийся с интерферон-мРНК;
6.Позитивные группы разбили на 8 подгрупп, содержащих по 64 клона, и вновь провели тестирование. Разбиение на подгруппы повторяли до тех пор, пока не идентифицировали клон, содержащий полноразмерную интерферон-кДНК человека.
Одним из первых рекомбинантных препаратов интерферона является Роферон-А (α-2a), «Hoffman la Roche» – высокоочищенный белок, который содержит 165 аминокислот с молекулярной массой около 19 кDа. Интерферон имеет четыре остатка цистеина, участвующих в образовании двух дисульфидных связей (Cys 1-Cys 98 и Cys 29-Cys 138), одна из которых (Cys 29-Cys 138) существенна для проявления интерфероном антивирусной активности. Препарат получают по технологии рекомбинантной ДНК
239
с использованием генно-инженерного штамма E. Coli, с включенной плаз- |
интерферона в культуральной жидкости разрешает очищать его по упро- |
||||
мидой, содержащей ген интерферона лейкоцитов человека. |
щенной схеме: отсутствие необходимости сбора биомассы, её дезинтегра- |
||||
Экспериментально показано, что уровень экспрессии генов интерфе- |
ции, денатурации белков с целью растворения «телец включения» и рена- |
||||
рона действительно увеличивается пропорционально числу тандемных ко- |
турации молекул интерферона. Отсутствие стадии концентрирования кле- |
||||
пий гена, по крайней мере, до четырех копий на плазмиду. Однако тандем- |
ток (сбор биомассы), а также и клеточных белков, разрешает получить вы- |
||||
ные повторы иногда оказываются нестабильными и со временем некото- |
сокоочищенный препарат интерферона менее сложным методом, чем тот |
||||
рые из них или даже все утрачиваются плазмидой. |
который используется при «плазмидной» технологии. Очищение осу- |
||||
Плазмидозависимые технологии являются высоко производитель- |
ществляется ионообменной хроматографией. По предложенной техноло- |
||||
ными, тем не менее, имеют серьезный недостаток – целевой продукт, как |
гии был получен рекомбинантный интерферон «Лаферон». В полученном |
||||
правило, образовывает в клетке нерастворимые в воде кристаллообразные |
препарате обнаруживается не менее 95 % белка интерферона. |
||||
формы, так называемого «тельца включения». Процесс изготовления и |
Первый ген интерферона был выделен в начале 80-х годов. С тех пор |
||||
очистки интерферона при использовании плазмидной технологии состоит |
обнаружено несколько различных интерферонов. Исходя из химических и |
||||
из таких стадий: собирание клеточной биомассы; её дезинтеграция; рас- |
биологических |
свойств, их |
можно подразделить |
на |
три группы: |
творение «телец включения» путем денатурации белковых структур с по- |
α-интерферон, β-интерферон, γ-интерферон. Интерферон-α и β-интерферон |
||||
мощью мочевины или гуанин-дихлорида; ренатурация денатурированных |
синтезируются клетками, обработанными препаратами вирусов или вирус- |
||||
молекул интерферона; очистка интерферона от балластных компонентов. |
ной РНК, а γ-интерферон вырабатывается в ответ на действие веществ, |
||||
Стадия ренатурации является ключевой стадией в процессе получения ре- |
стимулирующих рост клеток. Интерферон-α кодируется семейством генов, |
||||
комбинантных белков, в процессе которой денатурированный белок при- |
включающих |
как минимум |
15 неаллельных генов, |
в |
то время как |
обретает нативную пространственную структуру, обеспечивающую его |
β-интерферон и γ-интерферон кодируются одним геном каждый. Подтипы |
||||
биологическую активность. В процессе ренатурации молекул интерферона |
α-интерферона проявляют разную специфичность. Например, при провер- |
||||
наблюдается образование некорректных внутренне- и межмолекулярных |
ке эффективности интерферона-α1 и интерферона-α2 на обработанной ви- |
||||
связей. Это приводит к частичному образованию неправильных мономер- |
русом линии клеток быка эти интерфероны проявляют сходную противо- |
||||
ных форм рекомбинантного интерферона, конформация которых отличает- |
вирусную активность, в случае же обработанных вирусом клеток человека |
||||
ся от таковой у естественного интерферона, и возникновению его олиго- |
интерферон-α2 оказывается в семь раз активнее, чем интерферон-α1. Если |
||||
мерных структур, которые отсутствуют в естественном интерфероне. |
противовирусная активность проверяется на клетках мыши, то интерфе- |
||||
Предложен оригинальный способ получения рекомбинантного ин- |
рон-α2 оказывается в 30 раз менее эффективным, чем интерферон-α1. |
||||
терферона с использованием встроенного в бактериофаг гена интерфе- |
Было предпринято несколько попыток создать интерфероны с ком- |
||||
рона. Бактериофаг, заражая бактериальную клетку, размножается в ней, |
бинированными свойствами, используя тот факт, что члены семейства |
||||
копируя многократно свою ДНК и встроенный в неё ген интерферона, син- |
α-интерферона различаются по степени и специфичности своей противо- |
||||
тезирует свои белки, в том числе и интерферон. На определенной стадии |
вирусной активности. Теоретически этого можно достичь, если соединить |
||||
развития бактериофаг лизирует бактериальную клетку. Интерферон выхо- |
части последовательности генов разных α-интерферонов. Это приведет к |
||||
дит в культуральную жидкость, причем, в водорастворимом состоянии, не |
образованию гибридного белка с другими свойствами, чем у каждого из |
||||
образовывая нерастворимых форм. Синтез организован таким образом, что |
исходных белков. Сравнение последовательностей кДНК интерферона-α1 |
||||
интерферон накапливается вне клетки (в культуральной среде), поэтому не |
и интерферона-α2 показало, что они содержат одинаковые сайты рестрик- |
||||
образовывает «телец включения», как это имеет место в плазмидной тех- |
ции в позициях 60, 92 и 150. После расщепления обеих кДНК в этих сайтах |
||||
нологии получения интерферона (Intron-A, Roferon-A и др.). Накопление |
и последующего лигирования фрагментов было получено несколько гиб- |
||||
240 |
|
|
241 |
|
|
ридных генов. Эти гены экспрессировали в E. coli, синтезированные белки очистили и исследовали их биологические функции. Проверка защитных свойств гибридных интерферонов на культуре клеток млекопитающих показала, что некоторые из них проявляют большую активность, чем родительские молекулы. Кроме того, многие гибридные интерфероны индуцировали образование 2′-5′-олигоизоаденилат-синтетазы в контрольных клетках. Этот фермент участвует в синтезе 2′-5′-связанных олигонуклеотидов, которые в свою очередь активируют латентную клеточную эндорибонуклеазу, расщепляющую вирусную мРНК. Другие гибридные интерфероны проявляли большую, чем родительские молекулы, антипролиферативную активность в культурах различных раковых клеток человека.
Кроме рекомбинантных α-интерферонов, которые относятся к конкретной разновидности этого типа интерферонов, начат также выпуск так называемых «консенсусных» интерферонов. К ним относятся, в частности, интерферон «Альфакон-1» или «Инферген» («Амген»). Интерфероны с такими последовательностями вообще не существуют в природе, а представляют собой новые заданные комбинации аминокислотных последовательностей известных субтипов. Такие комбинированные интерфероны являются более эффективными, чем рекомбинантные интерфероны на основе природных субтипов.
Однако генно-инженерные интерфероны представлены белками только одной определенной формы, которая не подвергается посттрансляционной модификации и не идентична природным интерферонам, что может ограничить биологическую активность соответствующей композиции. Intron (α-2b) –Schering Plough, Roferon A (α-2a) – Hoffman la Roche, Лафе-
рон (α-2b) – Украина представляют собой исключительно (α-2)-форму интерферона. Интерфероны, полученные с помощью природных источников, таких как лимфобластоидная клеточная линия человека или лейкоциты периферической крови человека, представлены многими формами. По имеющимся данным препараты полученные из природных источников могут содержать более 15 белков с молекулярной массой от 18 до 25 кDа, обладающих антивирусной, антиростовой и иммуннорегуляторной активностью.
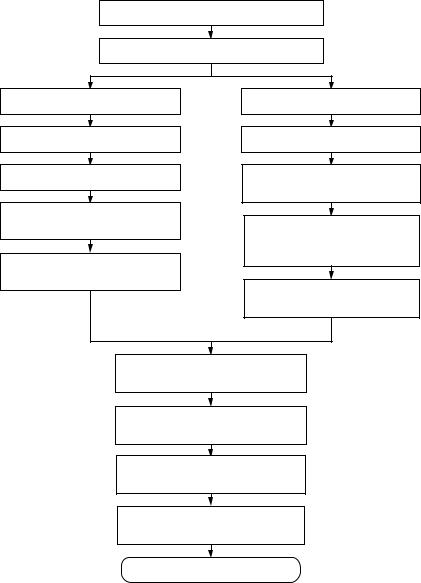
Технология получения рекомбинантного интерферона представлена на рис. 18.
242
Приготовление питательных сред
Получение посевного материала
Плазмидо-зависимый вариант
Производственное культивирование
Сбор биомассы и ее лизис
Выделение и очистка телец включения
Растворение телец включения и ренатурация интерферона
Фаго-зависимый вариант
Производственное культивирование
Отделение биомассы от культуральной жидкости
Концентрирование и очистка культуральной жидкости ультрафильтрацией
Стерилизующая фильтрация раствора интерферона
Очистка интерферона ионообменной хроматографией
Очистка интерферона гель-хроматографией
Ультрафильтрация раствора интерферона
Стерилизующая фильтрация субстанции интерферона
Контроль субстанции интерферона
Рисунок 18 – Технология получения рекомбинантного интерферона
243
За последнее десятилетие создано значительное количество штам- |
насыщения до и 10 % после индукции лактозой. Культивирование продол- |
|
мов-продуцентов рекомбинантного интерферона, в частности, интерферо- |
жали до тех пор, пока прирост оптической плотности культуры за 1 час не |
|
на-α-2b. Получение гена интерферона-α-2b также проводится различными |
снижался до 1 ОЕ. Клетки дрожжей после ферментации центрифугировали |
|
методами, определяемыми конкретным исследователем. В качестве при- |
20 минут при 7000 об/мин и 4 °С. |
|
мера приводим краткую технологию получения интерферона-α-2b, пред- |
2. Приготовление экстракта клеток, содержащего интерфе- |
|
ложенную российскими учеными. |
рон-α-2b. |
|
E. Coli BL21(DE3) – хорошо известный лабораторный штамм микро- |
Клетки штамма E. Coli ресуспендировали в 0,5 л буфера для дезинте- |
|
организма, используемый для получения рекомбинантных продуктов, |
грации (50 мМ натрия фосфорнокислого, 1 мМ ЭДТА, 100 мМ натрия хло- |
|
например, используя данный штамм, был получен продуцент гормона ро- |
ристого, рН – 6,8). Клетки разрушали на проточном дезинтеграторе, ис- |
|
ста человека (соматотропин). Ген интерферона-α-2b был собран из олиго- |
пользуя 2 цикла на каждую партию. Тельца включения собирали центри- |
|
нуклеотидов методом ПЦР по теоретической последовательности и клони- |
фугированием в течение 30 минут при 10000 об/мин и температуре 4 °С. |
|
рован по сайтам BamHI и NdeI в экспрессионном векторе pET22(b+)-2Tф. |
Осадок телец включения ресуспендировали в 0,5 л отмывочного буфера |
|
Штамм BL21(DE3) был трансформирован полученной плазмидой |
(50 мМ натрия фосфорнокислого, 1 мМ ЭДТА, рН – 7,2) и собирали цен- |
|
pET22(b+)-IFN-α-2b-2Tф. После отбора клонов, содержащих вставку гена |
трифугированием, дважды повторяя эту процедуру. |
|
интерферона, методом рестрикционного анализа целевая структура гена |
Как видно из приведенных данных после дезинтеграции клеток оса- |
|
интерферона была подтверждена путем определения нуклеотидной после- |
док телец включения подвергали двукратной отмывке. Это необходимо |
|
довательности ДНК. |
для удаления основной массы нуклеиновых кислот и части растворимых |
|
Технология получения интерферона-α-2b представлена следующими |
белков. Подобные этапы отмывки присутствуют почти во всех работах с |
|
стадиями: |
тельцами включения, причем встречаются как относительно простые про- |
|
1. Культивирование штамма продуцента интерферона-α-2b. |
цедуры с использованием буфера, в котором проводили лизис или дезинте- |
|
Единичную колонию штамма E. Coli BL21(DE3)[pET22(b+)-IFN-α- |
грацию, так и многоступенчатые, с использованием 3–8 М мочевины, три- |
|
2b-2Tф] пересевали в колбу с 30 мл среды Лурия – Бертани, состоящей из |
тона Х-100 и других неионных детергентов. По данным авторов, при пер- |
|
триптона – 10 г/л, дрожжевого экстракта – 5 г/л, натрия хлористого – |
вой отмывке из осадка телец включения уходит около 30 % клеточного |
|
10 г/л, ампициллина – 100 мкг/мл. Культивирование проводили в течение |
белка от начального количества, а при второй – около 8 %. Потери интер- |
|
7 часов при температуре 37 °С и перемешивании 240 об/мин. Далее выра- |
ферона по данным SDS-ПААГ-электрофореза при этом незначительны – не |
|
щенный инокулят переносили в лабораторный ферментер (рабочий объем |
более 2 %. |
|
1,5 л), используя синтетическую среду, содержащую: триптон – 10 г/л, |
Осадок отмытых телец включения переносят в емкость с частично |
|
дрожжевой экстракт – 5 г/л, глюкозу – 30 г/л, ампициллин – 200 мкг/мл. |
растворенными навесками буфера для растворения (29 мМ трис-HCl, |
|
Поддерживали рН культивирования на уровне 6,8 с помощью 12 %-ного |
рН – 8,0, 1 мМ ЭДТА, 7 М гуанидин-HCl). Доводят общий объем до 150 мл |
|
раствора гидроксида аммония. В качестве индуктора в ростовую среду до- |
и совместно растворяют твердые вещества в течение 2–4 часов при посто- |
|
бавляли лактозу до концентрации 20 г/л или изопропил- -D-тиогалактозид |
янном перемешивании и подогреве до комнатной температуры. В раствор |
|
добавляют навеску сульфита натрия (до 30 г/л) и проводят реакцию окис- |
||
до конечной концентрации 1 мМ. Аэрация составляла 25 % растворенного |
||
кислорода от насыщения до и после индукции изопропил- -D- |
лительного сульфитолиза в течение 12 часов при слабом перемешивании и |
|
комнатной температуре. |
||
тиогалактозидом. Аэрация составляла 25 % растворенного кислорода от |
||
244 |
245 |
Как известно, в тельцах включения интерферон присутствует в виде высокомолекулярных полимеров (вплоть до 5 млн. Da) с хаотично замкнутыми S-S связями. В связи с этим после растворения осадка телец включения в гуанидин-гидрохлориде проводили реакцию окислительного сульфитолиза, которая переводит полимеры интерферона в мономерную форму. Эта реакция более эффективна, чем восстановление S-S связей дитиотреитолом или 2-меркаптоэтанолом. Кроме того, полное растворение белков из телец включения достигается при более высоких концентрациях белка в растворе гуанидин-гидрохлорида, чем при использовании восстановителей.
Полученный раствор S-сульфоната интерферона разводили до 1,5 л 50 мМ Na-фосфатным буфером, осветляли центрифугированием и обессоливали на колонке сефадекса G-25 (11,5 х 55 см), уравновешенной 50 мМ Na-фосфатным буфером, рН – 8,0. На выходе получают 2,5 л раствора интерферона.
В полученный обессоленный и освобожденный от гуанидина и сульфита раствор добавляли 2-меркаптоэтанол до конечной концентрации 1,2 мМ и инкубировали 48 часов при 6 °С без перемешивания. Далее в ренатурирующую смесь медленно добавляли 2 М уксусную кислоту до рН 4,5. Выпавший осадок удаляли центрифугированием. Раствор ренатурированного интерферона концентрировали с помощью ультрафильтрации в 5–10 раз на ультрафильтрационной ячейке Biomax 10 кDa.
После проведения обессоливания и восстановления 2-меркапто- этанолом интерферон подвергали ренатурации в течение двух суток, а затем изменяли рН с 8,0 до 4,5. На данном этапе образуется осадок, который удаляли центрифугированием. В осадок выпадает часть бактериальных белков, а также часть неправильно собранного интерферона. На этом этапе возможны существенные потери интерферона, количество которого зависит от условий хранения телец включения (замораживались ли они и на какой срок, или же сразу после получения растворялись в гуанидингидрохлориде), а также от содержания примесей нуклеиновых кислот и бактериальных белков в ренатурирующей смеси, которые могут являться центром коагуляции интерферона.
246
3. Ионно-обменная хроматография.
Ренатурированный интерферон-α-2b наносят на колонку SP-Toyopearl 550 C (2,6 х 100 см), уравновешенную 10 объемами 50 мМ Na-ацетатного буфера, рН – 4,5, со скоростью потока 600 мл/ч. Колонку промывали тем же буфером, пока оптическая плотность при 280 нм не снижалась до базовых значений. Далее интерферон элюировали 50 мМ Na-ацетатным буфером, рН – 4,5 в градиенте натрия хлористого от 0 до 400 мМ (25 объемов колонки) со скоростью потока 250 мл/ч. Фракции анализировали ВЭЖХ, объединяя те из них, содержание мономера нативного интерферона в которых составляло более 80 %.
Несомненный интерес представляют данные сравнения влияния индукторов интерферона – лактозы и изопропил- -D-тиогалактозида. Показано, что при добавлении лактозы накопление интерферона в клетках происходит медленнее, чем при добавлении изопропил- -D-тиогалактозида. Однако культура при длительной ферментации достигает большей оптической плотности при использовании лактозы. Более того, при последующем выделении интерферона из образующихся в обоих случаях телец включения, субстанция интерферона, полученная из биомассы, индуцированной лактозой, содержит меньшее количество модифицированного (ацетилированного, содержащего N-концевой метионин или окисленные аминокислотные остатки) интерферона по сравнению с препаратом, полученным при индукции изопропил- -D-тиогалактозидом.
Приведенные методы получения интерферонов описывают выделение рекомбинантного продукта из нерастворимых агрегатов – телец включения. Однако, в ряде случаев, например, при получении дельтаферона, который синтезируется в растворенном виде, возможно использование надосадочной жидкости после отделения штамма-продуцента. В качестве штамма-продуцента использовали E. Coli MH-1 трансформированные плазмидой pIFN-дельта10. На первой стадии очистки дельтаферона использовали хроматографию на КМ-сефарозе. Дельтаферон практически полностью сорбируется на колонке при значении рН – 8,3; затем элюируется с сорбента в градиенте хлористого натрия от 0 до 0,5 М и при значении рН от 8,3 до 6,5. Доочистку дельтаферона проводили с помощью повторной хроматографии на КМ-сефарозе. Наносимый раствор белка дово-
247