Материал: Билеты по физе (с ответами)
3.Общее периферическое сопротивление сосудов (ОПСС) - сопротивление сосудов току крови, возникающее вследствие вязкости крови, трения о стенки сосудов и вихревых движений
4.Артериальным пульсом называются ритмические колебания артериальных стенок, обусловленные прохождением пульсовой волны. Пульсовая волна это расширение артерий в результате систолического повышения артериального давления. Пульсовая волна возникает в аорте во время систолы, когда в нее выбрасывается систолический порция крови и ее стенка растягивается. Обычно определяют его следующие параметры:
1. Частота пульса. В норме 60-80уд/мин.
2. Ритмичность. Если интервалы между пульсовыми волнами одинаковы пульс ритмичный.
3. Скорость пульса. Это быстрота пульсового повышения и понижения давления. При патологии может наблюдаться быстрый или медленный пульс.
4. Напряжение пульса. Определяется силой, которую необходимо приложить для того, чтобы пульс прекратился. Например, при артериальной гипертензии наблюдается напряженный пульс.
5. Наполнение. Складывается из высоты пульсовой волны и частично напряжения пульса. Зависит от величины систолического объема крови. Если сила сокращений левого желудочка падает, пульс становятся слабым.
5.Объективное исследование пульсовой волны осуществляют с помощью сфигмографии.
Это метод графической регистрации пульса. Сфигмография позволяет рассчитать такие физиологические показатели, как скорость распространения пульсовой волны, упругость и эластическое сопротивление артериального русла, а также диагностировать некоторые заболевания сердца и сосудов.
Сфигмограмма (СФГ) – запись пульсовых колебаний артерии. Сфигмографический датчик, преобразующий механические колебания в электрические, устанавливается на поверхности кожи над пульсирующей артерией. Восходящая часть кривой СФГ (анакрота) возникает за счет систолического выброса крови из левого желудочка; нисходящая часть кривой (катакрота) – за счет диастолического оттока крови в дистальном направлении. Инцизура на нисходящей части кривой совпадает с моментом захлопывания аортальных клапанов. Кровь, отраженная от аортальных клапанов, оттекает в дистальном направлении, вызывая появление еще одного (дикротического) подъема на кривой СФГ.
Билет 12
1. Рецепторы, их классификация, механизмы возбуждения первичных и вторичных рецепторов. Адаптация рецепторов, ее механизмы.
Рецептор служит для восприятия раздражений и их первичного анализа. Восприятие раздражения осуществляется путем трансформации энергии раздражения в нервный импульс. Различают первичные и вторичные рецепторы. В первичном рецепторе воспринимающая зона представляет собой окончание дендрита чувствительного нейрона; во вторичном рецепторе воспринимающей зоной является специальная рецепторная клетка, синаптически связанная с окончанием дендрита чувствительного нейрона. Первичные: все кожные рецепторы и рецепторы внутренних органов, проприорецепторы, рецепторы обоняния, термо- и хеморецепторы ЦНС. Вторичные: вкусовые, фоно-, фото-, вестибулорецепторы. В первичных рецепторах – рецепторный потенциал – потенциал действия. Во вторичных – рецепторный потенциал – выделение медиатора – генераторный потенциал – потенциал действия
Адаптационные процессы в рецепторах могут определяться внешними и внутренними факторами. В качестве внешнего фактора в механизме адаптации могут выступать свойства вспомогательных структур.
Внутренние факторы механизма адаптации связаны с изменениями физико-химических процессов в самом рецепторе.
2. Эритроциты, физико-химическая характеристика, функции. Понятие об эритроне. Возрастные и половые изменения количества эритроцитов. Механизм регуляции количества эритроцитов. Гемолиз эритроцитов его виды, значение.
Красные кровяные тельца (эритроциты) – безъядерные высокоспециализированные клетки организма, обеспечивающих транспорт: - кислорода, связывается с гемоглобином; вна форма транспортировки углекислого газа – соли угольной кислоты (бикарбонаты);- углекислого газа, который связывается с гемоглобином, благодаря карбоангідразній реакции в эритроцитах образуется осно - много других веществ, которые адсорбируются на поверхности эритроцитов (например питательные вещества). Количество эритроцитов в единице объема крови составляет: - у мужчин: 3,9 – 5,5 x 1012/л; - у женщин: 3,7 – 4,9 x 1012/л. Форма эритроцитов (двояковігнуті диски) обеспечивает максимальную площадь поверхности каждой клетки и наименьшее расстояние диффузии от поверхности к центру клетки. Диаметр эритроцитов – 7,5 мкм, но они способны к прохождению через капилляры меньшего диаметра, благодаря своей способности к деформации. Эритроциты не имеют ядра и митохондрий, их энергетический обмен проходит анаэробным путем (без использования кислорода) – все эти приспособления направлены на обеспечение транспортировки кислорода. Особенностью углеводного обмена является образование в эритроцитах 2,3-дифосфогліцеролу (2,3-ДФГ), который уменьшает сродство гемоглобина к кислороду (улучшает диссоциацию оксигемоглобина и отдачу кислорода тканям).
Механизмы регуляции количества эритроцитов в крови: повышается способность крови транспортировать кислород. Такие изменения возникают при любом стрессе и при физической нагрузке. быстрое увеличение количества эритроцитов в единице объема циркулирующей крови (ОЦК) выход эритроцитов из депо (венозные сосуды и селезенка) сужение большинства сосудов, в том числе и венозных, которые депонируют эритроциты 1. Нервные механизмы регуляции обеспечивают быструю смену количества эритроцитов в единице объема крови за счет их перераспределения между депо и активной циркуляцией. Главным механизмом является активация симпатического отдела вегетативной нервной системы (а точнее – симпато-адреналовой системы – САС)
Возрастные изменения:
Так, количество эритроцитов
(красные кровяные клетки) у новорожденного составляет 4,3-7,6 млн. на 1 мм3 крови, к 6 месяцам количество эритроцитов снижается до 3,5-4,8 млн. на 1 мм3, у детей 1 года – до 3,6-4,9 млн. на 1 мм3 и в 13-15 лет достигает уровня взрослого человека. Надо подчеркнуть, что содержание форменных элементов крови имеет и половые особенности, например, количество эритроцитов у мужчин составляет 4,0-5,1 млн. на 1 мм3, а у женщин – 3,7-4,7 млн. на 1 мм3.
Осуществление эритроцитами дыхательной функции связано с наличием в них гемоглобина
, являющегося переносчиком кислорода. Содержание гемоглобина в крови измеряется либо в абсолютных величинах, либо в процентах. За 100% принято наличие 16,7 г гемоглобина в 100 мл крови. У взрослого человека обычно в крови содержится 60-80% гемоглобина. Причем содержание гемоглобина в крови мужчин составляет 80-100%, а у женщин – 70-80%. Содержание гемоглобина зависит от количества эритроцитов в крови, питания, пребывания на свежем воздухе и других причин.
Содержание гемоглобина в крови также меняется с возрастом. В крови новорожденных количество гемоглобина может варьировать от 110% до 140%. К 5-6-му дню жизни этот показатель снижается. К 6 месяцам количество гемоглобина составляет 70-80%. Затем к 3-4 годам количество гемоглобина несколько увеличивается (70-85%), в 6-7 лет отмечается замедление в нарастании содержания гемоглобина, с 8-летнего возраста вновь нарастает количество гемоглобина и к 13-15 годам составляет 70-90%, т. Е. достигает показателя взрослого человека. Снижение числа эритроцитов ниже 3 млн. и количества гемоглобина ниже 60% свидетельствует о наличии анемического состояния (малокровия).
Гуморальные механизмы регуляции происходят посредством еритропоетинів, которые стимулируют созревание эритроцитов (эритропоэз) и их выход из красного костного мозга в кровь.
Данные механизмы регуляции обеспечивают медленное повышение количества эритроцитов в ОЦК. Еритропоетини образуются из глобулинов плазмы крови под влиянием еритрогенів. Еритрогени образуются в почках при таких условиях: а) уменьшение количества кислорода в крови (гипоксия); б) уменьшение кровоснабжения почек. Механизм: гипоксия – выработка эритрогенов почками – стимуляция образования эритропоетинов – их влияние на красный костный мозг – тусиление эритропоэза – увеличение количества эритроцитов в ОЦК – увеличение транспорта кислорода кровью – уменьшение гипоксии .
Гемолиз (гр. haima-кровь + lisis-растворение) - процесс разрушения оболочки эритроцитов и выход гемоглобина в плазму крови. Гемоглобин эритроцитов, выходя в плазму крови, окрашивает ее в красный цвет и кровь становится прозрачной - "лаковая кровь". По механизму происхождения различают несколько видов гемолиза.
-
Осмотический гемолиз может возникнуть в гипотонической среде (содержание солей в нем меньше 0,85 %).
-
Химический гемолиз может быть вызван хлороформом, эфиром, растворами кислот и щелочей, некоторыми лекарственными веществами (например, сульфаниламидами), разрушающими белково-липидную оболочку эритроцитов.
-
Биологический гемолиз могут вызываться гемолизинами растительного происхождения (отравление грибами).
-
Температурный (или термический гемолиз) возникает при замораживании/размораживании крови в результате разрушения оболочки эритроцитов кристалликами льда. А при несчастных случаях замерзание крови в дистальных отделах конечностей.
-
Механический гемолиз происходит при сильных механических воздействиях на кровь, например, при встряхивании ампулы с кровью, при длительной циркуляции крови в системе аппаратов искусственного кровообращения.
-
Физиологический гемолиз – это процесс, постоянно протекающий в организме, в результате которого в селезенке происходит захват из кровотока и разрушение «старых» эритроцитов макрофагами.
3.Механизм регуляции всасывания белков , жиров , жирорастворимых витаминов и углеводов в различных отделах жкт. Механизм возникновения голода и насыщения.
Всасывание – это процесс транспорта веществ из полости пищеварительного тракта во внутренние среды организма (кровь и лимфу). Большинство веществ всасывается в кровь (продукты гидролиза белков, углеводов и вода), в лимфу всасываются продукты гидролиза липидов. Интенсивно проходят процессы всасывания в верхних отделах тонкого кишечника. Всасывание в желудке. Объем всасывания на данном отрезке пищеварительного канала очень мал. Здесь всасываются моносахариды, некоторые соли, вода и алкоголь. Всасывание в тонком кишечнике. Способность к всасыванию практически одинакова во всех отделах тонкого кишечника. У здорового человека всасывание питательных веществ, солей и воды проходит в верхних отделах тонкого кишечника, а нижние отделы являются резервом всасывания. У взрослого человека при нормальном питании за сутки всасывается около 100 г белков в виде аминокислот, 100 г жиров в виде глицерина и жирных кислот, 500 г углеводов в виде моносахаридов. Общие механизмы всасывания: Преобладают активные механизмы транспорта (с затратами энергии против градиента концентрации). Так всасываются много ионов, большинство аминокислот и моносахаридов. Существует и пассивный транспорт. Так в эпителиоциты из кишечника поступают жирные кислоты. Вода всасывается также пассивно по механизму осмоса, или с помощью ультрафильтрации. Улучшают всасывание следующие условия: - pH=7,4 - осмотическое давление, что = 7,6 Атм. - гидростатическое давление химуса, что = 6 – 8 см.вод.ст. Всасывание проводят ворсинки, в состав которых входят кровеносные сосуды и лимфатические капилляры. В состав ворсинок также входят гладком'язеві клетки, которые при всасывании ритмично сокращаются и этим вызывают периодическое уменьшение или увеличение объема и площади поверхности ворсинок, что усиливает всасывание. Механизм всасывания ионов Na+: В базолатеральних отделах мембран эпителиоцитов работают Na-насосы, активно (с затратой энергии АТФ) транспортируют ионы Na + из эпителиоцитов в интерстиций (межклеточную жидкость), а потом он пассивно поступает в кровь. За счет работы натриевых насосов в эпителиоцитах создается низкая концентрация Na + и вследствие этого он по градиенту концентрации пассивно входит в эпителиоциты из полости кишечника, т. е. в конечном результате всасывания Na проходит по механизму первичного пассивного активного транспорта. За сутки всасывается 25 – 35г. Na, главным образом в тонком кишечнике, хотя этот процесс проходит также и в толстом кишечнике. Механизм всасывания глюкозы: Углеводы всасываются только в виде моносахарина, преимущественно по механизму вторичного активного транспорта в комплексе с ионами Na. Na - насос с пзатратами энергии АТФ создает градиент концентрации ионов Na. На апикальной мембране есть белки-переносчики, которые имеют 2 активных центра. Один для связывания ионов Na, второй – для связывания моносахарина (например, глюкозы). Комплекс белок-переносчик – ион Na – глюкоза, движется к внутренней поверхности мембраны клетки, это движение вызывает градиент концентрации ионов Na в клетке и в полости кишки (этот градиент создается с помощью Na-го насоса действие которого было упомянуто выше). На внутренней поверхности мембран клеток комплекс распадается и в цитоплазму поступают ионы Na и глюкоза. Далее ионы Na удаляются из клетки Na-насосом, а глюкоза переходит в кровь пассивно по механизму диффузии. Белок-переносчик становится свободным и цикл повторяется снова. Механизм всасывания белков: Белки всасываются в основном в виде аминокислот (АК) по механизму активного транспорта вместе с ионами Na. Выделяют 5 белков-переносчиков для различных АК. Небольшая часть белков проходит в эпителиоциты в виде полипептидов по механизму третичного активного транспорта также в комплексе с ионами Na. В эпителиоцитах эти полипептиды гидролизуются до АК, которые дальше пассивно поступают в кровь. У маленьких детей возможно всасывание по механизму піноцитоза (мікровезикулярний транспорт), который имеет для детей большое значение, так как обеспечивает поступление в организм ребенка продуктов гидролиза молока. Механизм всасывания жиров имеет следующие особенности: 1) жиры всасываются преимущественно в виде жирных кислот и глицерина, хотя могут всасываться и моноглицериды; 2) жирные кислоты с длинными цепями и глицерин всасываются только в комплексе с желчными кислотами; 3) поскольку жиры плохо растворяются в воде, то они транспорта-ются к эпителиоцитам в комплексе с желчными кислотами. Желчные кислоты вместе с липидами образуют мицеллы цилиндрической формы. Внутри мицеллы размещается жирная кислота. Мицеллы свободно перемещаются в воде и подходят к мембранам эпителиоцитов. Здесь мицеллы распадаются на жирные кислоты и глицерин, которые поступают в клетку пассивно по механизму диффузии; 4) в эпителиоцитах проходит ресинтез нейтральных жиров (три-глицеридов), которые характерны для данного организма из жирных кислот и глицерина, поступивших с пищей; образуются хіломікрони, которые увеличивают водорастворимость жира;5) синтезирован нейтральный жир в эпителиоцитах соединяется с белками 6) хіломікрони транспортируются преимущественно в лимфу.
Билет 13
Пищевой центр возбуждается под влиянием комплекса различных факторов. их можно разделить на две группы: метаболиты крови и состояние пищеварительного тракта. Одним из механизмов, вызывающих чувство голода, является сокращение пустого желудка, которое воспринимается механорецепторами стенки желудка. Это важный, но далеко не единственный фактор, поскольку после денервации желудка или удаления его чувство голода сохраняется. Чувство голода также зависит от концентрации в крови некоторых веществ. Согласно так называемой глюкостатичною теорией, чувство голода наступает вследствие снижения в крови содержания глюкозы. Снижение его сказывается на глюкорецепторы гипоталамуса, синокаротиднои зоны и др.. Согласно другой теории, чувство голода обусловлено снижением в крови концентрации аминокислот, продуктов обмена липидов и других веществ. Чувство насыщения связано с раздражением рецепторов органов пищеварения, в частности желудка и двенадцатиперстной кишки. Особенно заметную роль играет их наполнения, при котором подавляется центр голода. Нервные влияния передаются посредством афферентов блуждающего и симпатического нервов. Гормон холецистокинин также уменшуе чувство голода. Различают два вида насыщения - сенсорное (первичное) и обменное (вторичное). Первичное насыщение возникает вследствие раздражения вкусовых, обонятельных рецепторов, механорецепторов рта и желудка. Оно возникает еще во время еды. В это время повышается концентрация в крови глюкозы, свободных жирных кислот, которые поступают из депо. Вторичное насыщения возникает несколько позже, только тогда, когда продукты гидролиза питательных веществ всасываются бы кровь и лимфу. В настоящее время некоторые гормоны (ХЦК-ПЗ, соматостатин, бомбезин, субстанция Р) усиливают насыщение и снижают чувство голода, наоборот, пентагастрин, инсулин, окситоцин
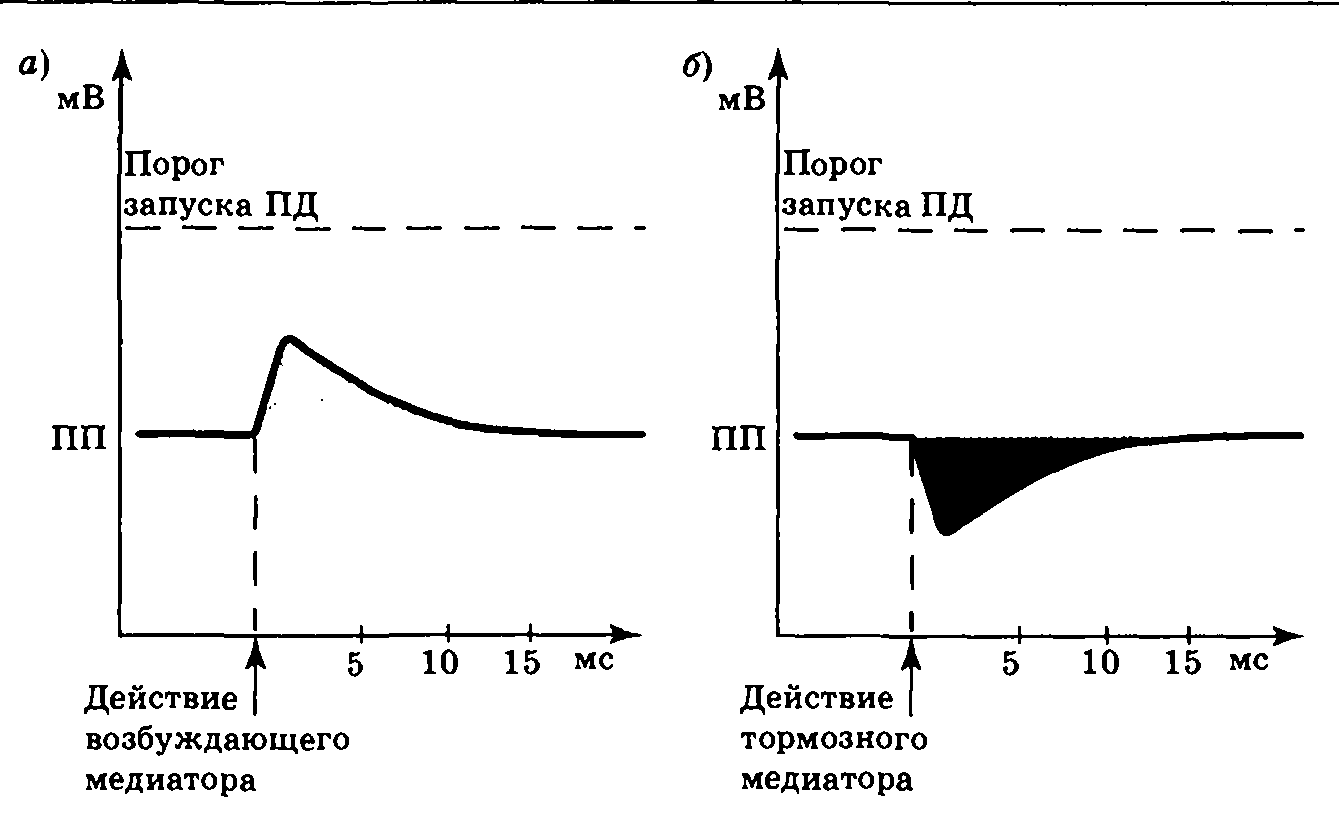
1.Торможение в центральной нервной системе. Его виды и физиологическая роль. Механизмы развития пре и постсинаптического торможения (Сеченов, Экклз) . Тормозные нейроны и их медиаторы. Тормозной постсинаптический потенциал (ТПСП). Механизм его возникновения. Нарисуйте схему ТПСП и сделайте необходимые обозначения.
Торможение- нервный процесс, что обусловлен возбуждением и внешне проявляется угнетением другого возбуждения
Торможение является важным фактором координационной деятельности ЦНС, участвует в обработке информации, поступающей к нейрону, выполняет охранительную роль.
Пресинаптическое торможение-связано с угнетением проведения нервных импульсов аксональных(пресинаптических) окончаниях. К возбужденному аксону подходит вставочный тормозной аксон что выделяет тормозной медиатор ГАМК
Постсинаптическое торможение- обусловлено выделением из пресинаптического окончания аксона тормозного медиатора, что понижает или тормозит возбудимостьсомы иди дендридов(аксо-сомальный, аско-дендритный синапс)
тормозные нейроны - клетки Реншоу в спинном мозге, грушевидные нейроны (клетки Пуркинье) коры мозжечка, звездчатые клетки коры мозга и др.
Медиаторами, или нейротрансмиттерами, нейронов ЦНС являются различные биологически активные вещества. В зависимости от химической природы их можно разделить на 4 группы:
1) амины (ацетилхолин, норадреналин, дофамин, серотонин),
2) аминокислоты (глицин, глутаминовая, аспарагиновая, гамма-аминомасляная - ГАМК)
3) пуриновые и нуклеотиды (АТФ);
4) нейропептиды (вещество Р, вазопрессин, опоидни пептиды и др.).
При действии тормозных нейромедиаторов в постсинаптической мембране открываются каналы для ионов хлора, вследствие чего ионы хлора входят в клетку, отрицательный заряд на внутренней стороне мембраны увеличивается и происходит гиперполяризация мембраны – образуется тормозной постсинаптический потенциал (ТПСП), который затрудняет образование ПД.
2.Фибринолиз, его механизмы. Антикоагулянтные механизмы. Значение в клинической практике.
Растворение кровяного сгустка такой же цепной ферментативный процесс, как и его образование. В настоящее время считается, что в крови постоянно протекают, как процессы образования небольших количеств фибрина так и процессы фибринолиза. В норме они уравновешены. И только в том вы случая, когда коагуляционная система дополнительно стимулируется продуктами повреждения сосудов и тромбоцитов, коагуляция начинает преобладать над фибринолизом.