Материал: Normalnaya_fiziologia_ekzamen2021
Диффузионная способность лёгких. Факторы, влияющие на газообмен через аэрогематический барьер (закон диффузии Фика).
Газообмен в легких осуществляются на основании диффузии
Диффузия - процесс газообмена через легочную мембрану в альвеолах.
Величина газообмена через легочную мембрану главным образом зависит от парциального давления каждого из газов. В то же время определенное значение имеют растворимость газов и температура. Газы диффундируют из области с более высоким парциальным давлением данного газа в область с более низким его давлением. Кислород поступает в кровь, диоксид углерода выводится из нее.
Для осуществления диффузии необходимы воздух, транспортирующий кислород в легкие, и кровь, утилизирующая этот кислород и выделяющая диоксид углерода. Воздух поступает в легкие во время легочной вентиляции, при этом осуществляется газообмен между ним и кровью.
Кровь из большинства участков тела возвращается по полой вене в легочную (правую) часть сердца. Из правого желудочка она выкачивается через легочную артерию в легкие, прокладывая себе путь к легочным капиллярам. Эти капилляры образуют плотную сеть вокруг альвеолярных мешочков. Легочные капилляры настолько малы, что эритроциты могут передвигаться по ним лишь в один ряд, подвергаясь воздействию окружающей легочной ткани. Здесь происходит процесс диффузии.
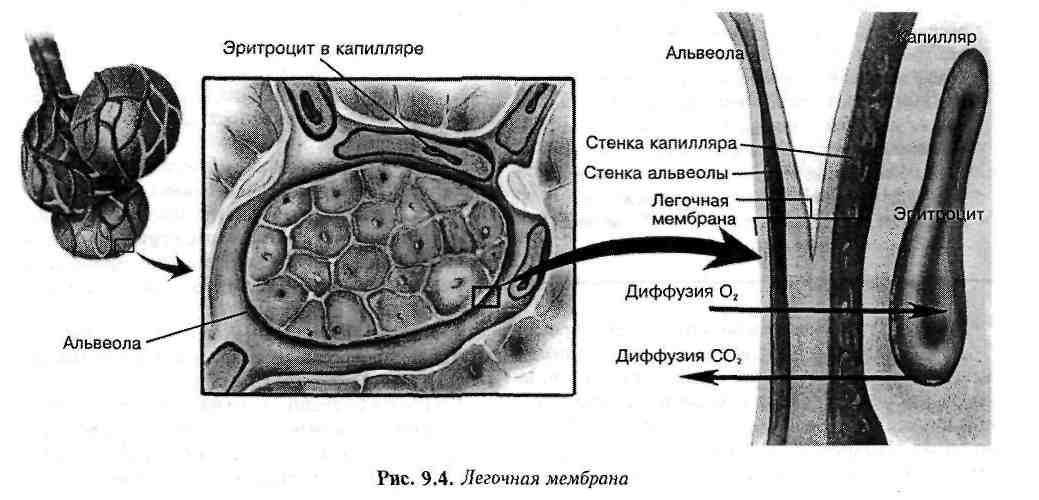
Альвеолярно-капиллярная мембрана ↑
Газообмен между воздухом в альвеолах и кровью в легочных капиллярах осуществляется через легочную мембрану (альвеолярно-капиллярную мембрану). Состоит из альвеолярной стенки; капиллярной стенки и их базальных мембран. Различие парциальных давлений газов в альвеолах и в крови создает градиент давления через легочную мембрану. Это является основой для осуществления газообмена во время диффузии кислорода и углекислого газа.
Диффузионная способность кислорода повышается при переходе из состояния покоя в состояние выполнения физической нагрузки. Когда организму требуется больше кислорода, процесс газообмена интенсифицируется.
Градиент давления обмена диоксида углерода меньше, чем градиент давления обмена кислорода, в то же время растворимость С02 в легочной мембране в 20 раз больше, чем растворимость кислорода, поэтому он легче диффундирует через нее даже при меньшем градиенте давления.
Скорость диффузии определяется множеством факторов:
-площадью диффузионной поверхности (А);
-толщиной мембраны (L);
-градиентом давления газов в альвеолярном воздухе и напряжения газов в крови (P1 - P2);
-коэффициентом диффузии (K).
1)площадь поверхности мембраны может значительно уменьшаться при воздействии многих факторов (удаление одного легкого уменьшает общую площадь дыхательной мембраны в 2 раза);
2)толщина мембраны может иногда увеличиваться (при появлении в интерстициальном пространстве отечной жидкости) (скорость диффузии газов при этом значительно снижается);
3)градиент давления определяет направление диффузии: если парциальное давление газа в альвеолах больше, чем его напряжение в крови, как это бывает с кислородом, диффузия совершается в направлении из альвеол в кровь;
4)величина коэффициента диффузии при переходе каждого газа через дыхательную мембрану находится в прямой зависимости от растворимости газа в мембране и в обратной зависимости от квадратного корня молекулярной массы этого газа;
Согласно закону диффузии (закон Фика) скорость диффузии

Транспорт кислорода кровью. Кривая диссоциации оксигемоглобина.
Большая часть кислорода в организме млекопитающих переносится кровью в виде химического соединения с гемоглобином. Свободно растворенного кислорода в крови всего 0,3%. Реакция оксигенации, превращение дезоксигемоглобина в оксигемоглобин, протекающая в эритроцитах капилляров легких:
Нв + 4О2 ⇄ Нв(О2)4
Реакция протекает очень быстро – время полунасыщения гемоглобина кислородом около 3 мс. Гемоглобин обладает двумя свойствами, которые позволяют ему быть идеальным переносчиком кислорода. Первое – способность присоединять кислород, второе – отдавать его. Способность гемоглобина присоединять и отдавать кислород зависит от напряжения кислорода в крови.
График, отражающий способность гемоглобина присоединять и отдавать кислород, - кривая диссоциации оксигемоглобина. По оси абсцисс на этом графике - количество оксигемоглобина в процентах ко всему гемоглобину крови, по оси ординат – напряжение кислорода в крови в мм рт. ст.
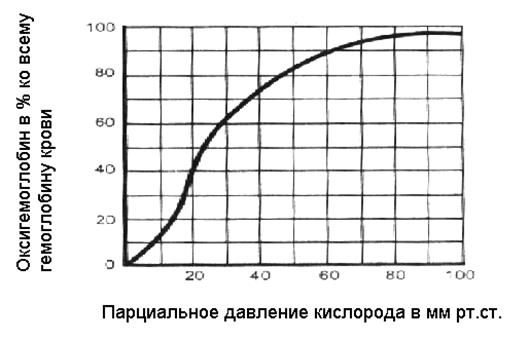
В соответствии с этапами транспорта кислорода: самая высокая точка соответствует тому напряжению кислорода, которое наблюдается в крови легочных капилляров – 100 мм рт.ст. (столько же, сколько и в альвеолярном воздухе). При таком напряжении весь гемоглобин переходит в форму оксигемоглобина – насыщается кислородом полностью. Один моль гемоглобина может связать 4 моля О2 , а 1 г Нв связывает 1,39 мл О2 в идеале, на практике 1,34 мл. Количество кислорода, которое может связать гемоглобин при условии его полного насыщения, - кислородная емкость крови (КЕК).
При снижении напряжения кислорода в крови до 60 мм рт.ст, насыщение практически не изменяется – почти весь гемоглобин присутствует в виде оксигемоглобина. Эта особенность позволяет связывать максимально возможное количество кислорода при снижении его содержания в окружающей среде.
Кривая диссоциации имеет s-образный характер, что связано с особенностями взаимодействия кислорода с гемоглобином. Молекула гемоглобина связывает поэтапно 4 молекулы кислорода. Связывание первой молекулы резко увеличивает связывающую способность, так же действуют вторая и третья молекулы. Этот эффект - кооперативное действие кислорода.
Артериальная кровь поступает в большой круг кровообращения и доставляется к тканям. Напряжение кислорода в тканях колеблется от 0 до 20 мм рт. ст., незначительное количество физически растворенного кислорода диффундирует в ткани, его напряжение в крови снижается. Снижение напряжения кислорода сопровождается диссоциацией оксигемоглобина и освобождением кислорода. Освободившийся из соединения кислород переходит в физически растворенную форму и может диффундировать в ткани по градиенту напряжения. На венозном конце капилляра напряжение кислорода равно 40 мм.рт.ст, что соответствует примерно 73% насыщения гемоглобина. Крутая часть кривой диссоциации соответствует напряжению кислорода обычному для тканей организма – 35 мм рт.ст и ниже.
Кривая диссоциации гемоглобина отражает способность гемоглобина присоединять кислород, если напряжение кислорода в крови высоко, и отдавать его при снижении напряжения кислорода.
Транспорт углекислого газа кровью. Химические реакции в эритроците при газообмене в легких и тканях (цикл Гендерсона).
Углекислый газ - продукт метаболизма клеток тканей, переносится кровью от тканей к легким. Выполняет жизненно важную роль в поддержании во внутренних средах организма уровня рН механизмами кислотно-основного равновесия. Транспорт углекислого газа кровью тесно взаимосвязан с этими механизмами.
В плазме крови небольшое количество углекислого газа находится в растворенном состоянии; при РС02= 40 мм рт. ст. переносится 2,5 мл/100 мл крови углекислого газа, или 5 %. Количество растворенного в плазме углекислого газа в линейной зависимости возрастает от уровня РС02.
В плазме крови углекислый газ реагирует с водой с образованием Н+ и HCO3. Увеличение напряжения углекислого газа в плазме крови вызывает уменьшение величины ее рН. Напряжение углекислого газа в плазме крови может быть изменено функцией внешнего дыхания, количество ионов водорода или рН — буферными системами крови и HCO3, например путем их выведения через почки с мочой. Величина рН плазмы крови зависит от соотношения концентрации растворенного в ней углекислого газа и ионов бикарбоната. В виде бикарбоната плазмой крови, в химически связанном состоянии, переносится основное количество углекислого газа — около 45 мл/100 мл крови (до 90%). Эритроцитами в виде карбаминового соединения с белками гемоглобина транспортируется примерно 2,5 мл/100 мл крови углекислого газа (5%). Транспорт углекислого газа кровью от тканей к легким в указанных формах не связан с явлением насыщения, как при транспорте кислорода, - чем больше образуется углекислого газа, тем большее его количество транспортируется от тканей к легким. Но между парциальным давлением углекислого газа в крови и количеством переносимого кровью углекислого газа имеется криволинейная зависимость: кривая диссоциации углекислого газа.
Многокомпонентная система взаимоотношений между структурами, участвующими в транспорте СО2 подчиняется закономерностям связанными с различной силой этих соединений, как кислот. Наиболее сильная - оксигемоглобин (ННbО2), затем угольная кислота (Н2СО3), наиболее слабая – дезоксигемоглобин (ННb).
Напряжение СО2 в тканях составляет 60 мм рт.ст., в протекающей артериальной крови примерно 40 мм рт.ст. В соответствии с диффузионным градиентом СО2 поступает из тканей в плазму капилляров.
Здесь некоторое количество (12%) остается в растворенном состоянии, большая часть соединяется с водой, образуя угольную кислоту. (Н2СО3).
В плазме крови эта реакция идет очень медленно. Мембрана эритроцита хорошо проницаема для углекислого газа, и он поступает внутрь эритроцита.
Здесь имеется фермент карбоангидраза, который при высоком Рсо2 ускоряет (в 10 тыс. раз – 2 мс) скорость этой реакции. Это первая реакция цикла Гендерсона.
СО2 + Н2 О→ Н2 СО3
Когда парциальное давление углекислого газа понижается (в легких) тот же самый фермент катализирует противоположную реакцию – разложение угольной кислоты на воду и углекислый газ. Образовавшаяся угольная кислота диссоциирует на ион бикарбоната (НСО3) и протон (Н+). Часть НСО-3 выходит в плазму, где соединяется с Na+ и предполагает компенсацию нарушенного равновесия энергетических зарядов.
Поскольку мембрана эритроцитов практически непроницаема для катионов, но сравнительно легко пропускает небольшие анионы, в обмен Н2 НСО3- в эритроцит поступают ионы Cl -.
Этот обменный процесс - хлоридный сдвиг (сдвиг Хамбурга). Протон Н+ присоединяется к дезоксигемоглобину.
Вторая реакция цикла Гендерсона связана с освобождением оксигемоглобина от кислорода (отдача кислорода) и образования дезоксигемоглобина. Оксигемоглобин транспортируется в виде калиевой соли.
КНbО2 → КНb + О2
Третья реакция цикла Гендерсона связана с тем, что угольная кислота (как более сильная, по сравнению с дезоксигемоглобином ННb) вытесняет (забирает) ион К+ из соединения КНb, оставшегося после диссоциации оксигемоглобина.
КНb + Н2 СО3 → КНСО3 + ННb
Четвертая реакция цикла Гендерсона связана с «судьбой» соединения КНСО3.
КНСО3 → К+ + Сl- → КСl
либо
КНСО3 → НСО3- + Na+ → NaHCO3
Часть этого соединения остается в эритроците; обеспечивая транспорт СО2. Часть диссоциирует на ион К+ который соединяясь с ионом Сl- (входящим из плазмы) образует КСl и ионы НСО-3, которые выходят из эритроцита в плазму, где соединяются с ионом Na+, образуя Na СО-3, одна из основных форм транспорта СО2.
События, происходящие в легких, сосудах малого круга кровообращения:
Первая реакция цикла Гендерсона. Напряжения О2 в легких составляет - 103 мм рт.ст., в венозной крови – 40 мм рт.ст. В соответствии с градиентом давлений кислород диффундирует в кровь, где образуется оксигемоглобин.
ННb + О2 → ННbО2
Вторая реакция цикла Гендерсона. Т.к. оксигемоглобин (ННbО2) более сильная кислота, чем угольная кислота, он вытесняет (связывает) ион К+ из бикарбоната.
КНСО3 + ННbО2 → КНbО2 + Н2СО3
Третья реакция цикла Гендерсона. Угольная кислота диссоциирует на Н2О и СО2. Углекислый газ диссоциирует по градиенту напряжений из венозной крови в альвеолы и выделяется (удаляется) из легких.
Н2СО3 → Н2О + СО2
Это происходит под действием карбоангидразы.
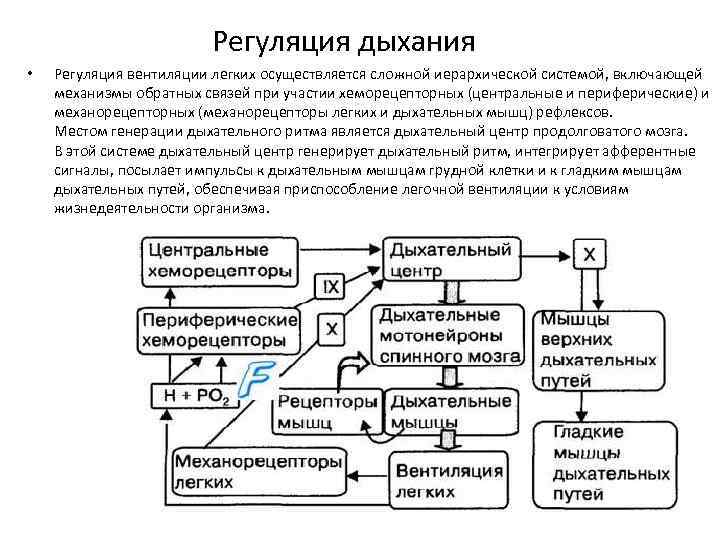
Структурно-функциональная организация дыхательного центра. Регуляция вентиляции легких.
Ритмическая последовательность вдоха и выдоха, изменение характера дыхательных движений в зависимости от состояния организма (покой, работа различной интенсивности, эмоциональные проявления и т. д.) обусловлены наличием дыхательного центра, расположенного в продолговатом мозге. Дыхательный центр - совокупность нейронов, обеспечивающих деятельность аппарата дыхания и его приспособление к изменяющимся условиям внешней и внутренней среды.
Дыхательный центр - парное, симметрично расположенное образование, в состав которого входят вдыхательная и выдыхательная части.
Предполагают, что в продолговатом мозге локализованы центры вдоха, выдоха и судорожного дыхания. В верхней части моста мозга (варолиев мост) находится пнеймотаксический центр, который контролирует деятельность расположенных ниже центров вдоха и выдоха и обеспечивает правильное чередование циклов дыхательных движений.
Дыхательный центр, расположенный в продолговатом мозге, посылает импульсы к мотонейронам спинного мозга, иннервирующим дыхательные мышцы. Диафрагма иннервируется аксонами мотонейронов, расположенных на уровне III-IV шейных сегментов спинного мозга. Мотонейроны, отростки которых образуют межреберные нервы, иннервирующие межреберные мышцы, расположены в передних рогах грудных сегментов спинного мозга (III-XII).
Дыхательный центр обладает автоматией. В нем автоматически возникает ритмическое возбуждение с частотой 14-16 раз в минуту из-за обменных процессов в самих нейронах, которые очень чувствительны к недостатку кислорода. Т.к. дыхательная мускулатура образована поперечнополосатой мышечной тканью, человек способен произвольно изменять вентиляцию легких. Это возможно, т.к. деятельность дыхательного центра продолговатого мозга находится под контролем коры больших полушарий.
В стенке легких имеются механорецепторы, которые возбуждаются при достижении легкими определенной степени растяжения при вдохе. От этих рецепторов поступают импульсы в дыхательный центр, взывая расслабление дыхательных мышц. В дыхательном центре ритмично возникает возбуждение и торможение.
Показатели и регуляция вентиляции легких в условиях измененной газовой среды (недостаток кислорода, избыток углекислого газа, изменённое атмосферное давление) и при физической нагрузке.
У тренированных людей при напряженной мышечной работе объем легочной вентиляции возрастает до 50—100 л/мин по сравнению с 5—8 л в состоянии относительного физиологического покоя. Повышение минутного объема дыхания при физической нагрузке связано с увеличением глубины и частоты дыхательных движений. У тренированных людей, в основном, изменяется глубина дыхания, у нетренированных — частота дыхательных движений.
При физической нагрузке увеличивается концентрация в крови и тканях углекислого газа и молочной кислоты, которые стимулируют нейроны дыхательного центра как гуморальным путем, так и за счет нервных импульсов, поступающих от сосудистых рефлексогенных зон. Активность нейронов дыхательного центра обеспечивается потоком нервных импульсов, поступающих от клеток коры головного мозга, обладающих высокой чувствительностью к недостатку кислорода и к избытку углекислого газа.
Одновременно возникают приспособительные реакции в сердечно-сосудистой системе. Увеличиваются частота и сила сердечных сокращений, повышается АД, расширяются сосуды работающих мышц и суживаются сосуды других областей.