Материал: Шандра О.А. Нормальна фізіологія. Вибрані лекції _ навч. посіб. _ О.А. Шандра, Н.В. Общіна _ О._ОГМУ, 2005. - 322 с
Ця сила, що діє в протилежному напрямку, зумовлена електричним зарядом аніонів Cl-. Коли К+, спрямований різницею залишкового тиску, залишає клітину, він виносить позитивний заряд, що за величиною відповідає негативному заряду на внутрішній стороні. При цьому виникає МПС. Полярність цього потенціалу така, що він перешкоджає виходу інших катіонів; позитивний заряд відштовхує позитивні іони. Вихід позитивних зарядів вже створює електричний потенціал, який перешкоджає виходу інших позитивних зарядів.
Отже, іон К+ забезпечує заряд зовнішньої поверхні мембран, який, відповідно до першого закону електрофізіології Германа, у спокої є позитивним. Так створюється різниця потенціалів між зовнішньою і внутрішньою поверхнею мембрани, утворюється МПС.
При збудженні МПС зменшується. Це зменшення МПС при збудженні тканини, від якої він відводиться, називається негативним коливанням потенціалу спокою, або потенціалом дії, який позначається ПД. Таке зменшення МПС можна пояснити за формулою: сила струму (J) прямо пропорційна різниці потенціалів (Е) і обернено пропорційна опо-
ру (R):
J= E/R.
Звідси випливає, що сила струму може зменшуватися або за рахунок зменшення різниці потенціалів, або за рахунок підвищення опору. Опір тканини під час збудження не тільки не збільшується, але й дещо зменшується. Значне зменшення сили струму відбувається за рахунок зменшення різниці потенціалів. Різниця потенціалів може зменшитися за рахунок збільшення негативного заряду всередині клітини або за рахунок зменшення позитивного заряду на зовнішній поверхні мембрани. Негативність усередині клітини максимальна і більше підвищуватися не може, тому необхідно зазначити, що відбувається зменшення позитивного заряду на поверхні мембрани і його зміна на негативний заряд. Отже, збуджена ділянка тканини електронегативна відносно електропозитивної, не збудженої в даний момент ділянки. Це і є ІІІ закон електрофізіології (ІІ закон: поверхня ушкодженої тканини має негативний заряд, І закон: зовнішня поверхня неушкодженої мембрани електропозитивна та ізопотенціальна).
Потенціал дії
Для реєстрації ПД використовують 2 відведення: однофазне і двофазне. При однофазному відведенні один електрод розташовують на поверхні клітини, а другий — всередині клітини або на її ушкодженому місці. При двофазному відведенні обидва електроди розміщуються на поверхні клітини. Відповідно до розташування електродів отриму-
14
ють однофазний або двофазний ПД. При одночасному розташуванні електродів спочатку реєструється МПС. При диполярному відведенні спочатку немає відмінності потенціалів, тобто неушкоджена і незбуджена тканина, відповідно до І закону електрофізіології, є ізопотенціальною. Після нанесення подразнення і появи збудження виникає негативність спочатку під одним електродом, а потім, оскільки збудження поширюється, — під другим. Реєструється двофазний ПД.
Величина ПД нервових клітин ссавців 100–110 мВ, скелетних і серцевих м’язових волокон — 110–120 мВ.
Тривалість ПД нервових клітин — 1–2 мс, у скелетних м’язах 5– 10 мс, у міокарді — понад 200 мс.
Природа ПД. Яким чином потенціал спокою, зазвичай підтримуваний на постійному рівні, порушується до такої міри, що виникає ПД
(рис. 3)?
Потенціал дії завжди виникає при деполяризації мембрани, тобто при втраті клітинною мембраною свого нормального заряду, приблизно до -50 мВ. Рівень потенціалу, при якому деполяризація дає початок потенціалу дії, називається порогом. При такому пороговому потенціалі заряд мембрани стає нестабільним; він порушується за допомогою внутрішнього механізму, який веде до реверсії полярності, швидкого наростання ПД до піка. Цей стан автоматичного прогресуючого порушення мембранного заряду називається збудженням. Звичайно збудження триває менше 1 мс. Воно подібне до вибуху — характеризується потужністю і швидким припиненням. Після фази деполяризації настає процес відновлення заряду мембрани, що відповідає стану спокою і називається реполяризацією.
Таким чином, ПД являє собою послідовну деполяризацію та реполяризацію мембрани — постійний для кожної клітини ауторегенеративний процес, який включається, як тільки рівень деполяризації мембрани перейде на пороговий потенціал. Кожен тип клітин має постійний і характерний для даного виду часові параметри ПД. Він практично не залежить від частоти збудження клітини. Оскільки форма ПД постійна, вважають, що збудження перебігає за законом «все або нічого».
Механізм ПД. Виникнення ПД пов’язане з короткочасним збільшенням проникності мембрани для іонів Na+ з подальшим підсиленням дифузії цих іонів за концентраційним градієнтом всередині клітини. Отже, якщо ПС або МП зумовлений іонами К+, то ПД — зумовлений іонами Na+, тому МП називається калієвим, а ПД — натрієвим. Дифузія іонів натрію всередину клітини призводить до зменшення МП. Зменшення МП до деякої критичної величини (звичайно на 10–30 мВ) викликає позитивний зворотний зв’язок, тобто зменшення МП нижче критичного рівня призводить до подальшого збільшення проникності мембрани для іонів Nа+, що супроводжується лавиноподібним підсиленням дифузії натрію всередину клітини. Посилений потік позитивно заряджених іонів
15
mV
0
І
КРД
-90
ІІ
b1
C
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
a1 |
|
|
|
|
|
c1 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|
d1 |
|
|
|
|
|
d2 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
a |
|
|
|
d |
|
|
|
|
b |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
с |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
a |
|
|
|
|
|
|
|
|
d |
|
|
|
|
|
|
|
e |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
c |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
b |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
A
B t
1
2
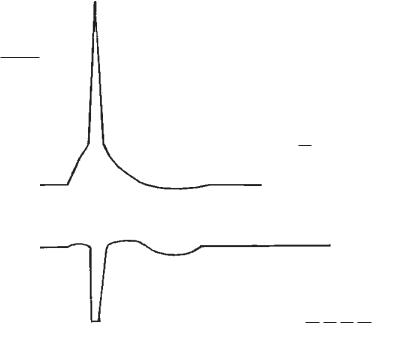
Рис. 3. Потенціал дії та зміни збудливості тканини під час збудження:
I — однофазний ПД на фоні мембранного потенціалу спокою; на осі ординат відкладено величини потенціалів в mV; на осі абсцис відкладено час в мс (t);
а— локальний потенціал відбиває деполяризацію мембрани під час локального збудження;
А — потенціал дії відбиває поширюване збудження:
а1 — швидка деполяризація мембрани за рахунок натрієвого струму усередину клітини через відкриті натрієві канали; b1 — момент інактивації натрієвих каналів; с1 — швидка реполяризація мембрани за рахунок роботи Na+/K+-на- соса; d1 — початок поширюваного збудження; d2 — кінець поширюваного збудження; d — тривалість ПД, тобто тривалість збудження;
В — низьковольтні потенціали:
а— локальний потенціал; b — слідова електронегативність; с — слідова електропозитивність;
С — овершут ПД.
II — зміни збудливості під час збудження:
1 — висхідний рівень збудливості визначають за порогом подразнення; 2 — нульовий рівень збудливості; а — невеличке збільшення збудливості під час локального збудження; b — абсолютна рефрактерна фаза; с — відносна рефрактерна фаза; d — підвищення збудливості (супернормальна фаза, «екзальтаційна» фаза за М. Є. Введенським); е — фаза субнормальної збудливості
16
Nа+ всередину клітини викликає спочатку зникнення надлишкового негативного заряду, а потім призводить до перезарядження мембрани: відбувається реверсія МП — зовнішня поверхня мембрани стає негативною по відношенню до внутрішньої. Настає фаза деполяризації, вона є наслідком виникнення короткочасного, спрямованого всередину натрієвого вхідного струму, що графічно відповідає за часом висхідній фазі ПД (рис. 3). Важлива роль Na+ в походженні ПД підтверджується експериментальними даними про те, що ПД можуть генеруватися тільки за високої зовнішньоклітинної концентрації Nа+. При нестачі зовнішньоклітинного Nа+ не може збільшуватися вхідний струм Nа+ незалежно від того, в якій мірі зростає К+ (градієнт Nа+), а отже не може виникнути деполяризаційна фаза ПД.
Другий етап виникнення ПД пов’язаний із більш тривалим періодом підвищеної проникності мембрани для іонів К+ і посиленням дифузії цих іонів з клітини назовні. Збільшення іонного потоку К+ призводить до зменшення проникності мембрани для іонів Na+ і зменшення їх потоку. Внаслідок цього потік Nа+ всередину клітини різко знижується, а для К+ зростає. Це призводить до появи більш тривалого вихідного струму, який відповідає фазі реполяризації, тобто відновлення полярності мембрани, і за часом відповідає низхідній фазі ПД (див. рис. 3). Отже, реполяризація мембрани відбувається не внаслідок зворотного переміщення іонів Nа+, а внаслідок виходу з клітини еквівалентної кількості іонів К+. Якщо підвищенню провідності для К+ запобігти деякими речовинами, наприклад тетраетиламонієм, мембрана після ПД реполяризується набагато повільніше. Це свідчить про те, що підвищення провідності для К+ є важливим фактором реполяризації мембрани.
Отже, ПД зумовлений циклічним процесом входу Nа+ в клітину і наступного виходу К+.
Сьогодні на молекулярному рівні інтенсивно вивчаються ті канали, якими рухаються іони Nа+ і К+ після появи ПД. Ці іонні канали специфічні й відрізняються від звичайних механізмів трансмембранного переносу іонів, які відомі з курсу біології та медичної фізики. Іонний канал складається з власне транспортної системи так званого ворітного механізму («воріт»).
Останніми роками увагу дослідників було прикуто до проблеми молекулярних механізмів, швидкої Nа+-системи. Розв’язанню цієї проблеми сприяло відкриття такої речовини, як тетродотоксин (синтезується в тканинах деяких риб і саламандр), який у концентраціях 10-10– 10-7 ммоль/л специфічно пригнічує або блокує швидку Nа+-систему. Молекула тетродотоксину блокує тільки вхід в Nа+-канал, причому незаблоковані канали залишаються інтактними. Зважаючи на це, можна підібрати концентрацію тетродотоксину, який блокує половину каналів,
17
і визначити кількість каналів на одиницю площі мембрани. Відповідно до таких розрахунків, існує приблизно 50 Nа+-каналів на 1 мкм мембрани, тобто канали розташовані в середньому на відстані 140 нм один від одного. Пора каналу відкривається тільки на короткий час, оскільки через 1 мс настає інактивація.
Ворота можуть знаходитися в двох положеннях: вони повністю закриті або ж повністю відкриті. Потенціал дії виникає тоді, коли у відповідь на підпорогову деполяризацію на короткий час відкриваються «ворота» для Nа+. Підвищення проникності для Na+ триває лише дуже короткий час. Потім проникність для Nа+ знову знижується. Процес, що веде до зниження раніше збільшеної натрієвої проникності, називається натрієвою інактивацією. Потім відразу ж відчиняються «ворота» для К+, які залишаються відкритими доти, доки не відновиться МПС
(рис. 4).
Властивості потенціалу дії. Першою властивістю ПД є те, що його амплітуда не залежить від сили подразнення, тобто від величини деполяризації. Якщо деполяризація досягає критичного рівня, то виникає ПД постійної величини незалежно від того, чи в подальшому підсилюватиметься подразнення і чи стане тривалішим. Ця властивість відома у фізіології у вигляді правила «все або нічого». Причиною такої властивості ПД є те, що він зумовлений клітинною реакцією, яка сама себе підтримує і має, як вважають, ауторегенеративний характер. Подразнення є тільки запускаючим моментом, поштовхом для внутрішньоклітинного механізму, який генерує ПД.
Другою властивістю ПД є рефрактерність. Викликати другий ПД відразу ж після виникнення першого неможливо. Якщо мембрана подразнюється відразу ж після ПД, збудження не виникає, ні при значенні подразнення, що відповідає порогу для попереднього ПД, ні за будь-якого сильнішого подразнення. Такий стан повної незбудливості, який у нервових клітинах триває 1 мс, називається абсолютним рефрактерним періодом. Після нього настає відносний рефрактерний період, коли шляхом значного подразнення можна все ще викликати ПД, хоча його амплітуда буде знижена порівняно з нормою. Рефрактерність зумовлена інактивацією Nа-системи під час попереднього ПД. Хоча при реполяризації мембрани стан інактивації минає, таке відновлення є поступовим процесом, що триває протягом кількох мілісекунд, впродовж яких Nа-система ще не здатна активізуватися.
Абсолютний рефрактерний період обмежує максимальну частоту генерації ПД. Наприклад, якщо абсолютний рефрактерний період завершується через 2 мс після початку ПД, клітина може збуджуватися з частотою максимум 500 в с, а якщо триває 500 мс, то 2 в с. З другої
18