Материал: Прищепа И.Н., Ефременко И.И. Нейрофизиология
Глава 1. ФИЛОГЕНЕЗ И ОНТОГЕНЕЗ НЕРВНОЙ СИСТЕМЫ
Этапы развития нервной системы
В процессе эволюции нервной системы происходило совершенствование, с одной стороны, ее отдельных элементов, в том числе нервных клеток, а с другой – ее общих физиологических свойств. Все это вместе обеспечивало расширение адаптационных возможностей организма. Поэтому выделяют три этапа в развитии нервной системы и, соответственно, три ее типа: диффузную – у кишечнополостных, узловую – у чле-
нистоногих и трубчатую – у позвоночных.
Диффузная, или сетевидная, нервная система. Первым этапом ее развития было формирование нервной системы диффузного типа, нервные клетки которой мало напоминают нейроны позвоночных. Ядро в них не дифференцировано, количество отростков невелико, их длина незначительна. Передача возбуждения от нейрона к нейрону осуществляется не только с помощью синапсов, но и через протоплазматические мостики. Нейроны слабо дифференцированы по функции. Скорость распространения возбуждения по нервным волокнам предельно низкая и составляет сотые и десятые доли метра в секунду. Основными особенностями нервной системы диффузного типа являются неопределенность связей, отсутствие четко выраженных входов и выходов, нена-
дежность функционирования.
Узловая нервная система. Нейроны узлового типа нервной системы коренным образом отличаются от элементов диффузного типа. На этом этапе филогенеза произошли количественные и качественные изменения нейронов – возросло их общее число и увеличилось количество вариаций нейронов, отличающихся по размеру, форме и числу отростков. Формирование нервных узлов привело к обособлению и структурной дифференцировке трех основных видов нейронов – афферентных (чувствительных), ассоциативных (вставочных) и эфферентных (двигательных).
В пределах нервной системы узлового типа (у насекомых, ракообразных) имеются также нервные волокна, покрытые многослойной оболочкой, которая напоминает миелиновую оболочку у нервных волокон позвоночных. Скорость проведе-
26
ния в этих волокнах намного выше, чем в аксонах такого же диаметра беспозвоночных, но меньше, чем у миелинизированных аксонов большинства позвоночных. Перехваты Ранвье здесь короче, чем у позвоночных. Именно этим обусловлена меньшая скорость распространения возбуждения по сравнению с аналогичным волокном у позвоночных. Как известно, механизм сальтаторного (скачкообразного) проведения возбуждения – довольно позднее эволюционное приобретение. Такой способ проведения возбуждения характерен лишь для позвоночных животных, имеющих нервную систе-
му трубчатого типа.
Трубчатая нервная система. Нервная система трубчатого типа – высший этап структурной и функциональной эволюции нервной системы. Все позвоночные имеют центральную нервную систему в виде нервной трубки, оканчивающейся головным мозгом. На уровне млекопитающих развивается кортиколизация – процесс формирования новой коры и экранных структур.
Центральная нервная система позвоночных состоит из спинного и головного мозга. Трубчатый вид имеет только спинной мозг. Головной мозг, развиваясь как передний отдел нервной трубки и проходя стадии мозговых пузырей, к моменту созревания претерпевает значительные конфигурационные изменения при существенном нарастании объема.
Спинной мозг при своей морфологической непрерывности
взначительной степени сохраняет принцип сегментарности и метамерности брюшной нервной цепочки узловой нервной системы. Так, простые сухожильные рефлексы сохраняются
вслучае целости только 2–3 сегментов спинного мозга, замыкающих эту рефлекторную дугу. С другой стороны, чрезвычайно развитый аппарат связей внутри спинномозговых путей обеспечивает целостность и разнообразие спинномозговых рефлексов. Среди них имеются сложные цепные рефлексы (шагательный), вовлекающие в процесс возбуждения практически весь спинной мозг. В процессе эволюции позвоночных животных вместе с прогрессирующим усложнением структуры и функции спинного мозга нарастает его зависимость от головного. Это хорошо видно на примере спинального шока, проявляющегося в торможении деятельности спинного мозга после отделения его от головного мозга. Такое торможение деятельности спинного мозга обусловлено в первую очередь прекращением нисходящих корригирующих влияний
27
со стороны головного мозга. Чем сильнее эти влияния, тем более глубокие и длительные последствия их прекращения.
Процесс цефализации, т.е. совершенствования структуры
ифункций головного мозга, у млекопитающих дополняется кортиколизацией – формированием коры больших полушарий
исовершенствованием ее функций. Если на уровне стволовых отделов и базальных ганглиев переднего мозга нейроны сгруппированы в виде отдельных специализированных ганглиев или ядер, то кора дает примеры совершенно новых принципов структурной и функциональной организации. Построенная по экранному принципу кора больших полушарий содержит не только специфические проекционные, но и значительные по площади ассоциативные зоны. Последние служат для корреляции различных сенсорных влияний, их интеграции с прошлым опытом для того, чтобы по моторным путям передать сформированные паттерны (рисунки) возбуждения и торможения для реализации поведенческих актов.
Вотличие от ганглионарных структур кора головного мозга обладает рядом свойств, характерных только для нее. Важнейшее из них – чрезвычайно высокая пластичность и надежность, как структурная, так и функциональная. Изучение этих свойств центральной нервной системы в эволюции позвоночных позволило обосновать вероятностно-статистический принцип организации высших функций мозга. Данный принцип в наиболее яркой форме выступает в коре головного мозга, являясь одним из приобретений прогрессивной эволюции.
Онтогенез нервной системы
Нервная система плода начинает развиваться на ранних этапах эмбриональной жизни. Из наружного зародышевого листка – эктодермы – по спинной поверхности туловища эмбриона образуется утолщение – нервная трубка. Головной конец этой трубки развивается в головной мозг, остальная часть – в спинной мозг.
У недельного эмбриона намечается незначительное утолщение в оральном (ротовом) отделе нервной трубки. На 3-й неделе зародышевого развития в головном отделе нервной трубки образуются три первичных мозговых пузыря (передний, средний и задний), из которых развиваются главные отделы головного мозга – конечный, средний, ромбовидный.
28
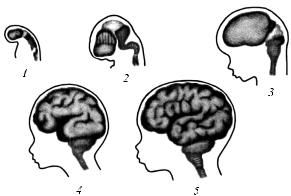
В последующем передний и задний мозговые пузыри расчленяются каждый на два отдела, в результате чего у 4–5-не- дельного эмбриона образуется пять мозговых пузырей: 1) конечный (телэнцефалон); 2) промежуточный (диэнцефалон); 3) средний (мезэнцефалон); 4) задний (метэнцефалон); 5) продолговатый (миелэнцефалон) (рис. 1). В последующем из конечного мозгового пузыря развиваются полушария головного мозга и подкорковые ядра; из промежуточного пузыря – промежуточный мозг (зрительные бугры, подбугорье); из среднего пузыря – средний мозг (четверохолмие, ножки мозга, сильвиев водопровод); из заднего пузыря – мост мозга (варолиев мост) и мозжечок; из продолговатого пузыря – продолговатый мозг. Задняя часть миелэнцефалона плавно переходит в спинной мозг.
Рис. 1. Последовательные стадии развития головного мозга из трех пузырей (1–5)
Из полостей мозговых пузырей и нервной трубки образуются желудочки головного мозга и канал спинного мозга. Полости заднего и продолговатого мозговых пузырей превращаются в IV желудочек; полость среднего мозгового пузыря – превращается в узкий канал (водопровод мозга, сильвиев водопровод), который сообщает между собой III и IV желудочки; полость промежуточного пузыря – в III желудочек, а полость конечного пузыря – в два боковых желудочка. Посредством парного межжелудочкового отверстия III желудочек сообщается с каждым боковым желудочком;
29
IV желудочек – с каналом спинного мозга. В желудочках
испинномозговом канале циркулирует спинномозговая жидкость.
Нейроны развивающейся нервной системы посредством своих отростков устанавливают связи между различными отделами головного и спинного мозга. Чувствительные нейроны заканчиваются рецепторами – периферическими окончаниями, воспринимающими раздражение; двигательные нейроны заканчиваются синапсами – контактными образованиями нервного волокна с мышцей.
К3-му месяцу внутриутробного развития выделяются основные части центральной нервной системы: большие полушария и ствол мозга, мозговые желудочки, а также спинной мозг. К 5-му месяцу дифференцируются основные борозды коры больших полушарий, однако кора остается еще недостаточно развитой. На 6-м месяце отчетливо выявляется функциональное превалирование высших отделов нервной системы плода над нижележащими отделами.
Головной мозг новорожденного имеет относительно большую величину, масса его в среднем составляет 1/8 массы тела, т.е. около 400 г, причем у мальчиков она несколько больше, чем у девочек. У новорожденного хорошо выражены борозды, крупные извилины, однако их глубина и высота невелики. Мелких борозд относительно мало, они появляются постепенно в течение первых лет жизни. К 9 месяцам первоначальная масса мозга удваивается и к концу первого года составляет 1/11–1/12 массы тела. К 3 годам масса головного мозга по сравнению с массой его при рождении утраивается, к 5 годам она составляет 1/13–1/14 массы тела. К 20 годам первоначальная масса мозга увеличивается в 4–5 раз и составляет у взрослого человека всего 1/40 массы тела. Рост мозга происходит в основном за счет миелинизации нервных волокон
иувеличения размера имеющихся уже при рождении примерно 20 млрд нервных клеток. Наряду с ростом головного мозга меняются пропорции черепа.
Мозговая ткань новорожденного малодифференцированна. Корковые клетки, подкорковые узлы, пирамидные пути недоразвиты, плохо дифференцируются на серое и белое вещество. Нервные клетки плода и новорожденного расположены концентрированно на поверхности больших полушарий и в белом веществе мозга. С увеличением поверхности головного мозга нервные клетки мигрируют в серое вещество; концен-
30