Материал: Прищепа И.Н., Ефременко И.И. Нейрофизиология
16.Основные характеристики безусловного рефлекса: а) приобретенный; б) врожденный; в) индивидуальный; г) видовой;
д) врожденно-обусловленное взаимодействие нейронов в рефлекторной дуге;
е) рефлекторная дуга формируется в процессе обучения.
17.Выделите фазы инстинктов: а) мотивации; б) поисковая; в) завершающая; г) запечатления.
18.Начальная, поисковая фаза проявления инстинкта запускается
с формирования в ЦНС:
а) мотивационного возбуждения; б) процессов запечатления; в) трансформации ритма; г) явления последействия.
19.Центры ЦНС, в которых возникает мотивационное возбуждение для проявления инстинктов, локализуются:
а) в коре больших полушарий; б) в коре мозжечка; в) в гипоталамусе;
г) в лимбической системе; д) в продолговатом мозге.
20.Укажите неправильный ответ. Формирование условных рефлексов происходит:
а) постоянно в течение жизни; б) при низком тонусе интегративных механизмов мозга;
в) на базе афферентной части врожденных безусловных рефлексов;
г) при активном состоянии коры («внимание»); д) благодаря образованию временных связей между нервными
центрами; е) на базе эфферентной части врожденных безусловных рефлексов.
21.Укажите неправильные ответы. Для формирования условных рефлексов необходимы следующие условия:
а) появление условного раздражителя должно предшествовать безусловному;
б) появление безусловного раздражителя должно предшествовать условному;
в) наличие сильного постороннего раздражителя;
206
г) активное состояние коры больших полушарий; д) нормальный тонус ретикулярной формации.
22.Укажите неправильные ответы. Для закрепления условных рефлексов необходимо:
а) многократное сочетание условного и безусловного раздражителей;
б) состояние мотивационного возбуждения центра безусловного рефлекса;
в) активное состояние коры больших полушарий; г) сниженный тонус ретикулярной формации мозгового ствола; д) действие постороннего раздражителя.
Глава 10. ТОРМОЖЕНИЕ В ЦНС
Торможение – это местный нервный процесс, приводящий к угнетению или предупреждению возбуждения. Одна из характерных черт тормозного процесса – отсутствие способности к активному распространению возбуждения по нервным структурам.
Впервые идею о том, что в ЦНС помимо возбуждения существует процесс торможения, высказал И.М. Сеченов. Исследуя рефлекторную деятельность лягушки с сохраненными зрительными буграми, И.М. Сеченов определял время сгибательного рефлекса – в ответ на погружение лапы животного
враствор кислоты происходило сгибание конечности в тазобедренном и коленном суставах. При помещении на зрительный бугор (таламус) лягушки кристаллов поваренной соли
вопытах И.М. Сеченова возникало торможение – удлинение времени сгибательного рефлекса. Это наблюдение позволило ученому прийти к выводу о наличии в ЦНС процесса тормо-
жения. В последующем такой вид торможения получил название сеченовское или центральное торможение.
|
С точки зрения физиологических механизмов, лежащих |
в |
основе торможения безусловных и условных рефлексов |
в |
ЦНС, в настоящее время принято различать первичное |
и вторичное торможение. Первичное торможение является результатом активации особых тормозящих структур (тормозящих нейронов), действующих на тормозимую клетку и вы-
зывающих в ней торможение как первичный процесс без предварительного возбуждения. Вторичное торможение воз-
никает в клетке без действия на нее специфических тормозя-
207
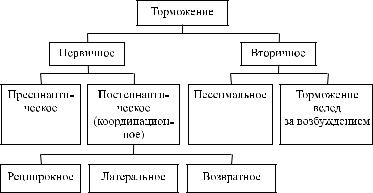
щих структур, как следствие ее возбуждения, т.е. вторично. К вторичному торможению относят пессимальное торможение и торможение вслед за возбуждением (рис. 30).
Рис. 30. Классификация торможения в ЦНС
Любой вид первичного торможения осуществляется с уча-
стием специализированных вставочных нейронов, или интернейронов, которые получили название тормозных нейронов.
Часть этих нейронов называются по имени автора, описавшего соответствующий тип (например, клетки Реншоу, клетки Уилкинсона, грушевидные клетки Пуркинье в мозжечке, звездчатые клетки коры и др.). Их аксоны образуют контакты непосредственно с тормозимой клеткой, либо аксоны тормозных клеток взаимодействуют с аксоном возбуждающего нейрона. Последний направляется к тормозимому нейрону, образуется аксо-аксональный синапс, который блокирует проведение возбуждения по аксону, и возбуждающий сигнал не доходит до тормозимого нейрона. Тормозные нейроны имеются во всех отделах ЦНС, но особенно их много в спинном мозге, мозжечке, базальных ядрах и в коре больших полушарий.
Виды торможения в ЦНС
Кпервичному торможению относятся пресинаптическое
ипостсинаптическое торможение. Разновидностью постсинаптического является реципрокное, возвратное и латеральное торможение.
208
Пресинаптическое, или фильтрационное, торможение
является частным случаем синаптических тормозных процессов, проявляющихся в подавлении активности тормозимого нейрона в результате уменьшения эффективности действия возбуждающих синапсов еще на пресинаптическом уровне. Оно развивается в пресинаптическом звене путем угнетения процесса высвобождения медиатора возбуждающими нервны-
ми окончаниями.
Постсинаптическое, или координационное, торможение развивается в основном в аксодендритных синапсах. Основой постсинаптического торможения является гиперполяризация постсинаптической мембраны тормозимого нейрона, представляющая собой тормозной постсинаптический потенциал (ТПСП). В качестве медиатора постсинаптического торможения могут выступать γ-аминомасляная кислота (ГАМК), глицин, норадреналин, дофамин, серотонин, эндозепины, эндогенные опиоиды, некоторые аминокислоты и пептиды. Все они приводят к повышению проницаемости постсинаптической мембраны тормозимого нейрона к ионам К+ и С1–, в результате чего возникает ТПСП. Важным условием эффективности тормозного процесса является наличие на тормозимом нейроне соответствующих рецепторов, способных воспри-
нять тормозной медиатор.
Реципрокное торможение было открыто английским физиологом Ч. Шеррингтоном и российским физиологом Н.Е. Введенским. Этот вид торможения основан на том, что одни и те же афферентные пути, через которые осуществляется возбуждение одной группы нервных клеток, обеспечивают через вставочные нейроны торможение других групп нейронов. Реципрокное торможение характерно как для спинного,
так и для головного мозга.
Возвратное, или антидромное, торможение наблюдается в отношении α-мотонейронов спинного мозга. При возбуждении α-мотонейрона нервный импульс направляется к мышечным волокнам, возбуждая их. Одновременно по коллатерали, идущей к тормозному нейрону (клетка Реншоу), импульс возбуждает тормозную клетку, которая, в свою очередь, вызывает торможение возбужденного ранее α-мотонейрона. Таким образом, α-мотонейрон, активируя, через систему тормозного нейрона сам себя затормаживает. Чем выше активность α-мотонейрона, тем выраженнее тормозное влияние клеток Реншоу на этот нейрон.
209
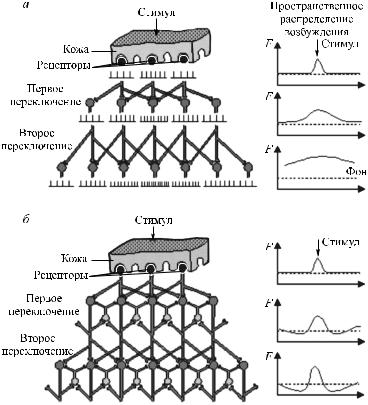
Латеральное торможение рассматривается как вариант возвратного торможения. Оно играет важную роль в организации обработки информации нейронами сетчатки. Суть его сводится к следующему. Под влиянием квантов света активируется фоторецептор. В свою очередь, это вызывает изменение активности биполярной клетки, связанной с данным фоторецептором. Одновременно активируется рядом расположенный тормозной нейрон, который блокирует проведение возбуждения от соседнего фоторецептора к ганглиозной клетке. Тем самым происходит «вытормаживание» информации в соседних участках. Таким способом создаются условия для четкого видения предмета (рис. 31).
Рис. 31. Эффект латерального торможения. Отрицательная обратная связь в процессе распространения возбуждения в нейронной сети (а, б)
210