Материал: Меледина Т.В., Давыденко С.Г. Дрожжи Saccharomyces cerevisiae. Морфология, химический состав, метаболизм
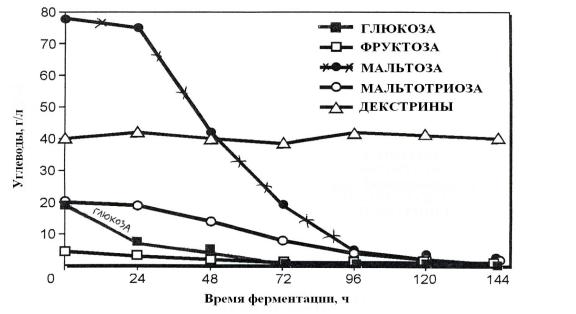
Рис. 6.2. Последовательность сбраживания сахаридов
6.2. ИСТОЧНИКИ АЗОТА
Содержание азота в клетках может достигать 10 % на СВ. Источниками азота для синтеза белка являются ионы аммония и аминокислоты. Однако рост клеток происходит быстрее, если в питательной среде содержатся аминокислоты, а не ионы аммония. Аминокислоты в процессе брожения потребляются последовательно. Это определяется свойствами и специфичностью пермеаз, локализованных в клеточной мембране дрожжей. Большинство поступающих в клетку аминокислот сначала подвергается действию трансаминаз, отщепляющих аминогруппы, при этом остается углеводный скелет, который далее метаболизируется.
Дрожжи не могут ассимилировать азот из органических соединений (кроме мочевины), нитратов и нитритов.
В дрожжевой промышленности и в виноделии в качестве источника азотного питания используют сульфат аммония (ГОСТ 10873–73), диаммонийфосфат марок А и Б (ГОСТ 8515–75) и раствор аммиака (ГОСТ 9–77). Очень редко в качестве источника азотного питания используется мочевина (ГОСТ 2081–75).
66
6.3. МИНЕРАЛЬНЫЕ КОМПОНЕНТЫ. МАКРО- И МИКРОЭЛЕМЕНТЫ
Активность некоторых ферментов может зависеть только от структуры самого белка, для других требуется также присутствие определенных групп небелковой природы – кофакторов. В роли кофакторов могут выступать ионы металлов или сложные органические соединения, называемые коферментами. Иногда для проявления активности фермента необходимо присутствие как тех, так и других.
Ниже приведены сведения о некоторых ферментах, коферментами в которых являются ионы металлов:
Zn+2 |
– кофермент алкогольдегидрогеназы, карбоксипептидазы; |
Mg+2 |
– кофермент пируватфосфокиназы, фосфогидролазы |
и фосфотрансферазы; |
|
Mn+2 |
– кофермент фосфотрансферазы; |
Fe+2 и Fe+3 – коферменты каталазы, пероксидазы и цитохромов; |
|
Cu+2 |
и Cu+ – коферменты тирозиназы и цитохромоксидазы; |
K+ – кофермент пируватфосфокиназы;
Na+ – кофермент плазматической АТФ-азы.
6.3.1.МАКРОЭЛЕМЕНТЫ
Кмакроэлементам относятся фосфор, калий, магний, натрий
исера.
Потребность дрожжей в фосфоре. Фосфор входит в состав нуклеиновых кислот, АТФ, фосфолипидов, полимеров клеточной стенки, в мембраны органоидов клеток. Он способствует поддержанию буферности, препятствующей сдвигу рН в цитоплазме клеток. Нехватка фосфора проявляется в замедлении скорости брожения и роста клеток.
Количество P2O5 в клетках определяется условиями культивирования, содержанием солей фосфора в среде и может достигать 5,5 % от СВ дрожжей.
Потребность дрожжей в калии. В хлебопекарных дрожжах содержание калия в пересчете на K2O в пределах от 1,4 до 4,3 % от АСБ, в пивных дрожжах – в среднем 2,4 % от СВ.
67
Калий активирует около 40 различных ферментов, в том числе оксидазы и дегидрогеназы. Ионы калия выполняют роль не только коферментов, но также входят в состав некоторых структур клетки,
вчастности, рибосом. Калий также участвует в регуляции транспорта ионов через клеточную стенку и через митохондриальную мембрану. В связи с увеличением скорости роста дрожжей потребность
вкалии увеличивается.
Ионы К+ определяют транспорт в клетки ортофосфата, играющего большую роль в энергетическом обмене дрожжей. Скорость этого процесса зависит как от вида микроорганизма, так и от условий культивирования. При энергетическом обмене, стимулируемом глюкозой, экзогенный калий поступает в клетку одновременно с фосфатом, при этом происходит отток ионов Н+ из дрожжей. Этот процесс является энергозависимым и происходит при участии фермента АТФ-азы.
Следует обратить внимание на обмен ионов К+ и Н+ через митохондриальную мембрану. В данном случае, помимо К+, в этом обмене участвуют также ионы Nа+ и ионы двухвалентных металлов Ca+2, Mn+2 и Sr+2. Переход этих ионов в митохондрию сопровождается выбросом эквивалентного количества Н+.
Для дрожжей экономический коэффициент по калию равен 54. Это означает, что 1 г элемента входит в состав 54 г АСБ.
Потребность дрожжей в магнии. Магний необходим как для энергетического, так и конструктивного обмена дрожжей, так как он является кофактором многих ферментов (см. раздел 5). Он стимулирует утилизацию мальтозы и мальтотриозы.
Экономический коэффициент в расчете на ионы потребленного магния составляет 540 г сухой биомассы на 1 г магния и обратно пропорционален количеству РНК в биомассе.
При составлении питательных сред следует учесть следующее соотношение между макроэлементами в пересчете на их оксиды: MgO:K2O:P2O5, которое равняется 1:10:20.
Потребность дрожжей в сере. Сера необходима, главным об-
разом, для синтеза серосодержащих аминокислот цистеина и метионина. Кроме того, небольшое количество серы требуется для образования сульфогрупп в некоторых коферментах, таких как биотин, кофермент А, липоевая кислота, тиамин и пиридоксин.
68
Было установлено, что в дрожжах при недостатке этого компонента в среде культивирования наблюдается снижение дыхательной активности клеток, как и при ингибировании роста дрожжей недостатком железа. Сера усваивается дрожжами из неорганических сульфатов, из серосодержащих аминокислот. Экономический коэффициент в расчете на ионы SО4-2 для пекарских дрожжей составляет 100 г сухой биомассы на 1 г.
6.3.2. МИКРОЭЛЕМЕНТЫ
Микроэлементы участвуют в метаболизме дрожжей и влияют на их химический состав, рост и размножение.
К микроэлементам, которые в первую очередь необходимы для роста дрожжей, относятся Zn, Cu, Ca, Mn, Fe, Co.
Элементы, редко требуемые для роста: B, Na, Al, Si, Cl, V, Cr,
Ni, As, Se, Mo, Sn, I. |
|
|
|
|
|
|
|
|
||||
|
Приблизительные значения |
экономического коэффициента |
||||||||||
прироста биомассы в расчете на |
некоторые |
элементы |
приведены |
|||||||||
в табл. 6.1. |
|
|
|
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
|
|
Таблица 6.1 |
|
|
|
Экономический коэффициент выхода биомассы |
|
|
|||||||
|
|
|
|
|
|
|
|
|
||||
|
|
Экономический коэффициент выхода биомассы , кг/г элемента |
||||||||||
|
Ca |
|
Fe |
|
Mn |
Zn |
|
Cu |
|
Co |
|
Mo |
|
1,0 |
|
6,7 |
|
20,0 |
20,0 |
|
100 |
|
100 |
|
100 |
В связи с тем что многие питательные ингредиенты среды также могут содержать микроэлементы и их количество не учитывается в расчете экономического коэффициента, данные, приведенные в табл. 6.1, приблизительны.
Потребность в микроэлементах может увеличиваться в несколько раз, когда культура испытывает стресс, например, при увеличении температуры выше оптимальной.
69
Микроэлементы могут накапливаться в клетках при культивировании их в среде с высокой концентрацией элементов. Так, биосорбция цинка в среде содержащей 0,8 мг Zn+2/мл, составляет
52,5 мг/г СВ.
Цинк входит в состав около 70 ферментов дрожжей. Он является кофактором ферментов клеточной стенки, стимулирует дыхательный метаболизм дрожжей, биосинтез белка, обмен нуклеиновых кислот, углеводов, вследствие этого он влияет на рост и размножение клеток. Ввиду участия цинка в высвобождении глюкозы (в виде гл-1-Ф) из гликогена увеличивается стрессоустойчивость дрожжей при их внесении в плотные питательные среды, в результате чего сокращается длительность лаг-фазы роста клеток.
В связи с тем что солодовое сусло, которое используется в технологии как хлебопекарных, так и пивных и спиртовых дрожжей, содержит недостаточное количество солей цинка, следует обратить внимание на факторы, влияющие на концентрацию этого микроэлемента в солоде, используемом для приготовления сусла. На содержание цинка в сусле влияют:
–степень растворения солода. Чем выше степень растворения солода, тем выше содержание цинка в горячем охмеленном сусле (без дробины);
–температура затирания. При 50 °С количество растворимого
всусле цинка больше, чем при затирании солода при 60 °С;
–гидромодуль затора. Жидкие заторы приводят к увеличению содержания цинка в сусле;
–рН затора. Биологическое подкисление способствует увеличению растворимости цинка.
Содержание ионов цинка в клетках должно быть на уровне 6 мг/100 г. Для получения этого количества в сусле должно содержаться не менее 0,08 мг цинка в литре. Ввиду того что цинк может аккумулироваться в клетках, дополнительное внесение этого элемента на этапе получения посевного материала гарантирует хорошие результаты при последующем размножении дрожжей в бедных питательных средах.
Расчет дополнительного внесения цинка можно сделать исходя из того, что экономический коэффициент для него составляет приблизительно 20 мг/г АСБ дрожжей (см. табл.6.1), или для прироста 100 г АСБ требуется 0,005 г цинка.
70