Материал: Меледина Т.В., Давыденко С.Г. Дрожжи Saccharomyces cerevisiae. Морфология, химический состав, метаболизм
действием литических ферментов. Если по какой-то причине (неблагоприятные условия культивирования) размер отделившейся дочерней клетки не достиг размера материнской, то длительность этой фазы увеличивается за счет времени, необходимого для достижения некоторого критического размера.
Реализовать стартовую программу могут только те клетки, которые достигли определенного размера, т.е. существует некий механизм, который регулирует начало клеточного деления, являющегося гарантией того, что только клетки, обладающие достаточными для завершения всего КЦ ресурсами, начнут последовательный ряд реакций, ведущих к делению клеток.
S – синтетическая фаза. Эта фаза заканчивается, когда почка достигает размера около 1/3 материнской. На этой стадии происходит удвоение ДНК.
G2 – постсинтетическая фаза. Происходит дальнейшее увеличение размера клетки. Идет распределение ядерного материала между материнской и дочерней клеткой.
М – митоз, включающий кариокинез (К) – деление ядра и цитокинез (Ц) – деление клетки. В период митоза размер почки продолжает увеличиваться вплоть до размера материнской.
Регуляция клеточного цикла
Регуляция клеточного цикла может осуществляться, по крайней мере, на двух различных уровнях:
первый – это цикл деления ДНК, который связан с репликацией ДНК и митозом;
второй – цикл роста, связанный с увеличением размера клетки. Обычно они функционируют вместе, поэтому средний размер
клеток любого штамма, при наличии некоторых колебаний, остается постоянным в широком диапазоне условий выращивания, за исключением стрессовых ситуаций.
Установлено, что в сахаромицетах размер клеток регулируется изменением длительности клеточного цикла, а не скорости роста. Причем изменения в клеточном цикле касаются, главным образом, продолжительности фазы G1.
Поведение популяции клеток в закрытой системе (периодическая культура) описывается кривой роста, включающей в себя несколько фаз.
6
Начальная фаза (лаг-фаза). Видимые признаки размножения дрожжей отсутствуют, однако обменные процессы в клетках протекают активно: синтезируются ферменты, необходимые для переноса
ирасщепления высокомолекулярных питательных веществ, внутри клеток синтезируются нуклеиновые кислоты и ферменты, необходимые для дальнейшего активного роста. За счет биосинтеза новых компонентов масса клеток увеличивается. Продолжительность фазы зависит от количества посевного материала, его возраста и физиологического состояния. Большое значение также имеет состав питательной среды и физико-химические параметры процесса (рН, температура, осмоляльность и т.п.). В этот период клетки наиболее уязвимы к различным стрессам.
Фаза ускорения роста – начало почкования, сопровождающееся дальнейшим увеличением количества биомассы в культуре.
Экспоненциальная (логарифмическая, лог-фаза). Характери-
зуется максимальной скоростью размножения. Численность клеток
иих биомасса возрастают в геометрической прогрессии. Из-за быстрого размножения возможно уменьшение размеров клеток.
Фаза замедления роста. В этот период концентрация питательных веществ в среде снижается, накапливаются токсичные продукты метаболизма, возрастает концентрация биомассы. В клетках происходит накопление резервных углеводов, в первую очередь гликогена. Это приводит к замедлению роста клеток и их размножения
идаже к частичной гибели клеток. Потребление кислорода постепенно снижается. Понижается активность дыхательных ферментов.
Стационарная фаза. В первой половине фазы прирост дрожжей продолжается с небольшой скоростью, при этом размеры клеток вновь увеличиваются. Оставшиеся в среде сбраживаемые углеводы используются клеткой в биосинтезе резервных углеводов (гликоген
итрегалоза).
Фаза затухания (отмирания) – период, когда в результате истощения питательной среды и максимального накопления продуктов обмена скорость отмирания и автолиза клеток превышает скорость их размножения. Автолитические процессы приводят к уменьшению общей численности клеток и изменению состава среды, в которой они находятся.
7
3. МОРФОЛОГИЯ ДРОЖЖЕЙ
Клетки S. сеrevisiae имеют округлую, яйцевидную или эллипсоидную форму; размер их колеблется от 2,5 до 10 мкм в поперечнике и от 4,5 до 21 мкм в длину. Размер и форма клеток одного и того же штамма определяются генетически и могут варьироваться в определенных пределах в зависимости от условий культивирования и последующих операций получения коммерческих дрожжей (обезвоживание).
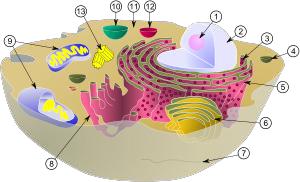
Клетки состоят из микроскопических (видимых в обычном микроскопе при увеличении в 600–900 раз) и субмикроскопических, видимых только в электронном микроскопе (увеличение от 15–20 тыс. раз), структур. Эти структуры можно подразделить на постоянно присутствующие и периодически обнаруживаемые в клетке. К первым относятся различные органеллы – клеточные структуры, выполняющие определенные функции. Это ядро с ядрышком, митохондрии, рибосомы, клеточная стенка, цитоплазматическая мембрана, эндоплазматический ретикулум (сеть), аппарат Гольджи, лизосомы, хитосомы, гликосомы и целый ряд других мембранных структур (рис. 3.1). Все клеточные органеллы окружены мембранами. В состав мембран входит большое количество фосфолипидов, причем их содержание как в количественном, так и в качественном составе определяется природой органеллы.
Рис. 3.1. Органеллы дрожжевой клетки:
1 – ядрышко; 2 – ядро; 3 – рибосома (маленькие точки); 4 – везикула;
5 – шероховатый эндоплазматический ретикулум (ER); 6 – аппарат Гольджи; 7 – цитоскелет; 8 – гладкий эндоплазматический ретикулум; 9 – митохондрия; 10 – вакуоль; 11 – цитоплазма; 12 – лизосома; 13 – центриоль и центросома
8
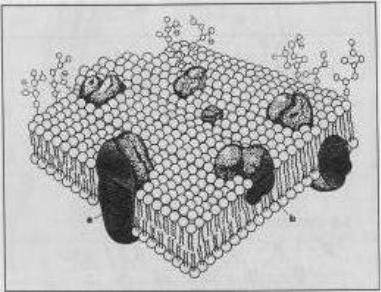
Мембраны органелл имеют трехслойную структуру. Они состоят из липидов, белков и небольшого количества углеводов. Липиды представлены в основном моно-, ди- и триглицеридами, глицерофосфатидами и стеролами – эргостеролом и зимостеролом. Каждая молекула фосфолипида состоит из гидрофобной, т. е. отталкивающей воду, и гидрофильной, притягивающей воду, частей. Гидрофильные части молекулы находятся на внешней стороне мембраны, а гидрофобные – на внутренней. Молекулы белка размещаются на поверхности мембраны или проникают внутрь нее (рис. 3.2).
Рис. 3.2. Модель клеточной мембраны (фосфолипиды и транспортные протеины)
Непостоянные структуры – включения – в отличие от органелл то возникают, то исчезают в процессе жизнедеятельности клеток. Будучи продуктами метаболизма клетки, включения отражают различные стороны и этапы ее физиологической активности. Этими непостоянными структурами являются внутриклеточные запасные соединения: жиры, гликоген и полифосфаты.
Включения могут быть представлены в виде более или менее плотных частиц – гранул, кристаллов или капель. Они обычно представляют собой скопления, видимые под микроскопом, либо без специальной обработки, либо после обработки различными красителями. Так, низкомолекулярные полифосфаты обнаруживаются в виде гра-
9
нул в вакуолях (волютин), жиры – в виде капель, гликоген выявляется при окрашивании клеток раствором Люголя.
3.1. КЛЕТОЧНАЯ СТЕНКА
Клеточная стенка является частью клеточной оболочки, в состав которой входит также периплазматическое пространство.
Клеточная стенка (КС) выполняет следующие основные функции:
1.Защита от воздействия окружающей среды.
2.Сохранение формы.
3.Участие в обменных процессах: регуляция поступления питательных веществ и выделение метаболитов.
4.Опосредованно участвует в процессах размножения.
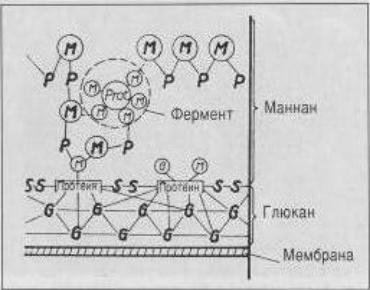
Клеточная стенка представляет собой слоистую структуру толщиной около 25 нм (рис. 3.3):
–1-й (наружный) слой – это тонкая липопротеиновая мембрана;
–2-й слой – значительно более толстый слой – представляет собой маннано-протеиновый комплекс;
–3-й слой состоит из глюкана, он имеет слоистую структуру.
Рис. 3.3. Модель клеточной стенки
10