Материал: физиология ответы
6.Корневое давление, физиологическая роль, зависимость от внутренних и внешних факторов.
Корневая система имеет поглощающую или всасывающую зону - это зона корневых волосков. Поступив в клетку корневого волоска вода становится частью живой системы - клетки растения - и подчиняется закономерностям, действующим в живой клетке. Передвижение по растению определяется двумя основными двигателями водного потока в растении:нижним двигателем водного потока или корневым давлением,верхним двигателем водного потока или присасывающим действием атмосферы.
Корневое давление создается при переходе воды из коры корня в сосудистую систему корня при прохождении воды через пропускные клетки перицикла, из которых вода под давлением как бы впрыскивается в сосуды ксилемы. Доказательством этого служат явления гуттации и "плача растений".
Гуттация - это выделение капельно-жидкой влаги листьями через гидатоды в условиях затрудненного испарения.
Плач растения - это вытекание пасоки (воды с растворенными в ней минеральными веществами, находящейся в ксилеме) из стеблей растений со срезанными побегами. Механизм образования корневого давления по-видимому состоит из двух аспектов:переноса воды по законам осмоса, дополнительной сократительной деятельности актомиозиновых белков, находящихся в перицикле и паренхимных клетках корня.
Присасывающее действие атмосферы определяется концентрацией водяных паров в атмосфере. Этот показатель в атмосфере почти всегда меньше, чем в листе растения, за исключением условий повышенной влажности воздуха, например, во время дождя, тумана.
Большая часть биоколлоидов клетки принадлежит к гидрофильным соединениям, способным к обратимым изменениям степени своей оводненности. Поглощая воду, коллоидная мицелла набухает, при отдаче же ею воды происходит отбухание. При этом в клетке развиваются весьма значительные силы, достигающие иногда сотен атмосфер.
Сила, которую нужно приложить к коллоидной системе, чтобы предотвратить поглощение ею воды, называется давлением набухания. Этому свойству биоколлоидов принадлежит важная роль в процессах поглощения протоплазмой воды, в передаче воды в вакуоль и в выделении воды клеткой.
При прорастании семян, например, обусловленном только явлением набухания, поглощение воды идет с силой до 105 килопаскаль.
Эндодерма - самый внутренний слой первичной коры, облекающий центральный цилиндр, часть эндодермы представлена тонкостенными клетками, называющимися пропускными. Именно через эти клетки вода под давлением проникает из клеток коры корня в центральный сосудистый цилиндр (ксилему).
Корневое давление зависит:
от условий влажности почвы (чем больше гидромодуль почвы, т.е. количество воды на единицу площади, тем интенсивнее идет поглощение воды растением),
от температуры почвы (ниже 12оС и выше 30оС поглощение воды замедляется),
от аэрации почвы (так как при нарушении аэрации ухудшается процесс дыхания, т.е. получения энергии клеткой, а, значит, и поглощения и передачи воды).
7. Транспирация: виды, механизмы, физиологическая роль и зависимость от внутренних и внешних факторов. Методы учета и возможности регулирования транспирации.
ТРАНСПИРАЦИЯ, у растений - потеря влаги в виде испарения воды с поверхности листьев или других частей растения.
Биологическое значение транспирации состоит в терморегуляции растения, в обеспечении деятельности верхнего концевого двигателя водного тока, при помощи которого осуществляется и поступление различных веществ, а также в регулировании насыщенности клеток водой, благодаря чему создаются оптимальные условия для процессов жизнедеятельности.
Типы транспирации:
1. Устьичная транспирация.
Число устьиц в листе в среднем составляет 50-500 на 1 кв. мм. Транспирация с поверхности листа идет почти с такой же скоростью как и с поверхности чистой воды. Это объясняется законом Стефана: через малые отверстия скорость диффузии газов пропорциональна не площади поверхности, а диаметру или длине окружности. То есть испарение из ряда мелких отверстий идет быстрее, чем из крупного отверстия того же диаметра. Это связано с явлением краевой диффузии. При диффузии из отверстий, стоящих на некотором расстоянии друг от друга молекулы воды, располагающиеся по краям, испаряются быстрее (рис. 3).
2. Кутикулярная транспирация.
Потеря воды растением через кутикулу имеет большое значение, когда устьица закрыты (засуха).
У молодых растений кутикулярная транспирация составляет около 50% всей транспирации, а у зрелых – 1/10. В старых листьях кутикулярная транспирация снова возрастает в результате разрушения и растрескивания кутикулы. Кутикулярна транспирация регулируется толщиной слоя кутикулы. Виды, имеющие толстый слой кутикулы, меньше теряют воду. А виды с тонкой кутикулой больше.
Одной из важных характеристик процесса является интенсивность транспирации — количество воды, испаряемое растением с единицы листовой поверхности в единицу времени. Высокая интенсивность транспирации, которую К. А. Тимирязев называл «необходимым злом», обусловлена тем, что атмосфера характеризуется очень низкими значениями водного потенциала
Как степень открытия устьиц влияет на интенсивность транспирации, так же внешние условия влияют на транспирацию , как на физический процесс.
Прежде всего на транспирацию влияет степень насыщенности атмосферы водой. Чем меньше относительная влажность воздуха, тем ниже его водный потенциал и тем быстрее идет транспирация. С увеличением влажности воздуха транспирация уменьшается; и при большой влажности воздуха происходит только гуттация
Транспирация зависит от температуры. С её повышением значительно увеличивается количество воды, насыщающей воздушное пространство, что увеличивает дефицит влаги в воздухе, а следовательно увеличивается и транспирация.
Свет – главный фактор, регулирующий транспирацию, который на испарение воды с водной поверхности практически не влияет. Влияние света на транспирацию связано прежде всего с тем, что клетки листа, содержание хлорофилл, поглощают не только инфракрасные лучи, но и видимы свет, необходимый для фотосинтеза. На фотосинтез лист расходует только 1-2% поглощенного света, а остальная энергия (98%) превращается в тепло, которая используется на испарение. Свет так же косвенно влияет на транспирацию, вызывая открытие устьиц.
В естественных условиях на интенсивность транспирации влияет ветер, но не так сильно как на испарение со свободной водной поверхности. В основном его влияние проявляется на третьем этапе устьичной транспирации, так как действие на внутренние ткани, из которых идет испарение, ограничено. Увеличение интенсивности транспирации происходит до 0,4 – 0,8 м/с, дальнейшее усиление ветра уже не влияет на транспирацию, так как в таком случае транспирация зависит от диффузии воды через устьица.
Транспирация зависит от влажности почвы. Здесь прямая зависимость. Чем меньше воды в почве, тем меньше её в клетках листа, а следовательно транспирация уменьшается., так как устьица прикрываются. Транспирация так же снижается, при недостатке недостаток кислорода в почве, что снижает дыхание корней, поглощение ими минеральных веществ, а следовательно и воды.
Интенсивность транспирации зависит от условий минерального питания, При недостатке азота, фосфора, или калия усиливает транспирацию. Транспирация сокращается если растение получает полное минеральное удобрение. Это объясняется тем, что поглощенные ионы и синтезированные вещества связывают воду, а испаряется только свободная вода.
Транспирация зависит и от внутренних условий.
Прежде всего интенсивность транспирации зависит от содержания воды в листьях. Всякое уменьшение воды в листьях уменьшает транспирацию.
Интенсивность транспирации зависит от количества свободной воды, водоудерживающей силы клеток. Чем меньше свободной воды, и меньше транспирация. Величина листовой поверхности так же является условием влияющим потери воды растением. Чем она больше, тем и больше теряется воды. Однако здесь могут быть различные приспособления растений, выработанных в процессе эволюции, снижающих испарение: опушенность листовой пластинки, погружение устьиц в мезофилл, толщина кутикулы.
Интенсивность транспирации зависит от соотношения поверхности корней и побегов. При увеличении доли корней транспирация увеличивается.
Возрастные изменения как листа, так и растения существенно влияют на интенсивность транспирации. Она может меняться у разных растений в зависимости от фаз и этапов онтогенезе.
8. Физиология устьичных движений . Значение устьиц в регулировании транспирации.
Устьица – это специфические образования, находящиеся в эпидермисе листа и предназначенные для газообмена листа с окружающей средой. Устьице состоит из 2 замыкающих клеток, между которыми находится устьичная щель. Замыкающие клетки, в отличие от всех остальных клеток эпидермиса, содержат хлоропласты.
Устьица способны к 2 типам движений: фотоактивному, которое индуцируется светом, и гидроактивному, которое индуцируется содержанием влаги в растении.
1. Фотоактивное движение устьиц
Обычно устьица открываются днем, когда идет фотосинтез и растению необходимо поглощать СО2, и закрываются ночью, чтобы растение не теряло лишнюю влагу.
2. Гидроактивное движение устьиц
Устьица во влажных условиях открываются, а при недостатке воды – закрываются
9. Транспирационный коэффициент и коэффициент водопотребления. Методы определения и величина у основных с/х культур.
●Транспирационный коэфф. – величина обратная продуктивной транспирации (кол-во созданного сух. в-ва на 1 л транспирир-ой воды). Он показывает, сколько воды растение затрачивает на построение единицы массы сухого в-ва. Варьирует от 100 до 500. У с/х культур значения близки, отличаются просовидные злаки (сорго, просо) – у них низкий.
Чем выше температура воздуха и ниже относительная его влажность, тем больше транспирационный коэффициент. Чем сильнее ветер, тем он больше. При сильном освещении – выше.
Определить трудно: листья в течение вегетации могут отмирать, учёт накопления массы корней еще более проблематичен. Определяют в вегетационных опытах: строгий учет кол-ва поливной воды и предотвращение испарения с корнеобитаемой среды.
●Коэффициент водопотребления (эвапотранспирационный) – отношение эвапотранспирации (суммарного расхода воды за вегетацию 1 га (исп. с почвы + транспирация)) к созданной биомассе/хозяйственно полезному урожаю. Для картофеля и пшеницы он в среднем около 300.
Можно рассчитать как разность в содержании влаги в метровом слое почвы в начале и конце вегетации плюс осадки и полив.
В засуху он выше (расход влаги больше, но продуктивность часто низкая). Увеличивается с повышением температуры. Чем более плодородна почва, тем ниже коэфф. водопотр. Коэфф. снижается при создании наиболее благоприятных условий.
11. Физиологические основы орошения с/х культур.
При избыточном орошении – ухудшение снабжение корней кислородом, уплотнение и засоление почва. При недостаточном – задержка роста листьев, снижение фотосинтеза.
Оптимальной является влажность почвы 70-80%.
Необходимо вычислять оросительную норму – кол-во воды, необходимое для полива определенной культуры за весь период вегетации в расчете на 1 га.
Для установления сроков орошения необходимо провести полив, когда растение ещё не испытывает недостатка в воде, но уже успело израсходовать почти всю воду с предыдущего полива.
Период наиб. чувствительности к недостатку влаги – критический период. Для установления времени полива определяют влажность почвы. Хороший показатель водоснабжения – движения устьиц: сразу после полива широко открыты, по мере исчерпания влаги закрываются. Также показателем является концентрация клеточного сока, которая возрастает при недостатке влаги.
Удобрение снижает количество необходимой растению воды, т. е. при оптимизации мин. питания оросительная норма ниже.
Глава 3 фотосинтез
-
Фотосинтез-основа биоэнергетики растений. Значение для обеспечения автотрофности.
Фотосинтез - это процесс образования органического вещества из углекислого газа и воды на свету при участии фотосинтетических пигментов. Главную роль в этом процессе играет использование энергии света для восстановления СО2 до уровня углеводов.
Растения – автотрофные организмы, они питаются теми органическими веществами, которые сами создают в процессе фотосинтеза. Образование органов у растения, их рост тесно связаны с синтезом органических веществ. Органические вещества служат для построения клеточных структур, откладываются в запас или вновь расщепляются в процессе дыхания до воды и углекислого газа.
Автотрофы — органифзмы, синтезирующие органические вещества из неорганических. Фототрофы - организмы, для которых источником энергии служит солнечный свет, они получают орг. в-во при помощи фотосинтеза.
Процесс фотосинтеза включает следующие этапы:
1) фотофизический;
2) фотохимический (световой);
3) ферментативный (темновой).
-
Общее уравнение фотосинтеза. Парциальные уравнения.
|
Общее уравнение: СO2 + 6H2O → C6H12O6 + O6 |
Суммарное уравнение: СO2 + H2O
|
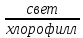 >
{СН2О} + О2
>
{СН2О} + О2Световая фаза (Н2О и О2)
Эта фаза происходит только в присутствии света в мембранах тилакоидов при участии хлорофилла, белков-переносчиков электронов и фермента — АТФ-синтетазы. Под действием кванта света электроны хлорофилла возбуждаются, покидают молекулу и попадают на внешнюю сторону мембраны тилакоида, которая в итоге заряжается отрицательно. Окисленные молекулы хлорофилла восстанавливаются, отбирая электроны у воды, находящейся во внутритилакоидном пространстве. Это приводит к распаду или фотолизу воды. Ионы гидроксила отдают свои электроны, превращаясь в реакционноспособные радикалы •ОН. Радикалы •ОН объединяются, образуя воду и свободный кислород.
Таким образом, в световую фазу происходит фотолиз воды, который сопровождается тремя важнейшими процессами: 1) синтезом АТФ; 2) образованием НАДФ·Н2; 3) образованием кислорода. Кислород диффундирует в атмосферу, АТФ и НАДФ·Н2 транспортируются в строму хлоропласта и участвуют в процессах темновой фазы.
Темновая фаза (СО2 и С6Н12О6)
Эта фаза протекает в строме хлоропласта. Для ее реакций не нужна энергия света, поэтому они происходят не только на свету, но и в темноте. Реакции темновой фазы представляют собой цепочку последовательных преобразований углекислого газа (поступает из воздуха), приводящую к образованию глюкозы и других органических веществ.
Первая реакция в этой цепочке — фиксация углекислого газа; акцептором углекислого газа является рибулёзодифосфат; катализирует реакцию фермент рибулёзодифосфат-карбоксилаза. В результате карбоксилирования рибулёзодисфосфата образуется неустойчивое соединение, которое сразу же распадается на две молекулы фосфоглицериновой кислоты. Затем происходит цикл реакций, в которых фосфоглицериновая кислота преобразуется в глюкозу. В этих реакциях используются энергии АТФ и НАДФ·Н2, образованных в световую фазу; цикл этих реакций получил название «цикл Кальвина».
6СО2 + 24Н+ АТФ → С6Н12О6 + 6Н2О.