|
1. |
Рибозо-5-фосфат |
НАДФ+ не відбувається (тобто гальмуються біо- |
|
|
|
|
|
|
|
Епімераза |
синтетичні процеси з участю НАДФ+), але, залеж- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
но від кількості еритрозо-4-фосфату, може відбува- |
|
|
Арабінозо-5-фосфат |
|
|
тися залучення метаболітів гліколізу до синтезу |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
2. Арабінозо-5-фосфат + Діоксіацетонфосфат |
пентозофосфатів, або, мабуть, при дуже складних |
|
умовах, пентозофосфати використовуються для |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Альдолаза |
утворення метаболітів гліколізу і підтримки енер- |
|
Октулозо-1,8-бісфосфат |
гозабезпечення в анаеробних умовах. |
|
|
|
|
|
|
|
|
|
|
|
Фосфатаза |
Клінічні аспекти пентозофосфатного шляху |
|
|
|
|
|
|
|
|
|
|
Октулозо-8-фосфат |
|
|
1. Пентозофосфатний шлях в еритроцитах по- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
3. Октулозо-8-фосфат + Еритрозо-8-фосфат |
стачає НАДФН для відновлення окисненого глу- |
|
татіону, ця реакція каталізується глутатіонре- |
|
|
|
|
|
|
|
|
Транскетолаза |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
дуктазою. Відновлений глутатіон руйнує в |
|
|
|
|
|
|
|
|
|
Фруктозо-6-фосфат + Глюкозо-6-фосфат |
еритроцитах Н2О2 за допомогою глутатіонперок- |
|
Схематично це можна виразити в такий |
сидази. Ця реакція має важливе значення, тому |
|
спосіб: |
|
|
|
|
|
|
|
|
|
|
|
що нагромадження Н2О2 може скоротити час |
|
|
|
|
Альдолаза |
життя еритроцитів (шляхом підвищення швид- |
|
|
|
|
кості окиснення гемоглобіну в метгемоглобін). |
|
|
С5 + С3 |
|
|
|
|
|
|
|
С8 |
|
|
|
|
|
|
|
|
|
G–S–S–G + НАДФН + Н+ |
|
|
|
|
|
|
|
|
|
|
Транскетолаза |
|
С8 |
+ С4 |
|
|
|
|
|
|
|
|
|
С6 + С6 |
|
Глутатіонредуктаза |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Біологічна роль пентозофосфатного шляху |
2 G–SH + НАДФ+ |
|
|
|
|
|
обміну вуглеводів |
2 G–SH + Н2О2 |
|
1. Пентозофосфатний шлях забезпечує орга- |
|
Глутатіонпероксидаза |
|
нізм пентозами, необхідними для біосинтезу нук- |
|
(селензалежна) |
|
G–S–S–G + 2 Н2О |
|
леотидів (наприклад, АТФ, ГТФ, ЦТФ, УТФ, |
|
НАД+, НАДФ+, ФАД, КоА та ін.). Фосфопенто- |
2. Недостатність глюкозо-6-фосфатдегідроге- |
|
зи необхідні також для біосинтезу нуклеїнових |
|
кислот — ДНК і РНК, де вони становлять близь- |
нази є причиною гемолітичної анемії за рахунок |
|
ко 40 % молекулярної маси нуклеїнових кислот. |
гемолізу еритроцитів. |
|
2. Пентозофосфатний шлях постачає віднов- |
У деяких груп людей спостерігаються мутації, |
|
лені форми НАДФ+ (НАДФН+Н+). Він задоволь- |
що спричинюють нестачу глюкозо-6-фосфатде- |
|
няє потреби організму в НАДФН+Н+ приблиз- |
гідрогенази, внаслідок чого порушується утво- |
|
но на 50 %. Відновлений НАДФ+ є донором |
рення НАДФН2, що призводить до гемолізу ери- |
|
Гідрогену для біосинтезу вищих жирних кислот, |
троцитів, особливо після прийому пацієнтами |
|
холестерину, |
пуринових основ та інших сполук, |
антималярійних препаратів, аспірину, сульф- |
|
використовується в мікросомальному окисненні |
аніламідів. Такі ж порушення виникають у па- |
|
при знешкодженні лікарських засобів та отрут у |
цієнтів при споживанні бобів Vicia faba — фавізм |
|
печінці, у синтезі жовчних кислот, стероїдних гор- |
(захворювання, на яке страждають мільйони лю- |
|
монів, у знешкодженні аміаку шляхом відновно- |
дей у країнах Африки й Азії). |
|
го амінування. Про важливу роль його для біо- |
|
|
|
синтезу вищих жирних кислот свідчить те, що у |
Метаболізм фруктози |
|
жировій тканині він становить приблизно 50 % |
|
від гліколізу (тобто 50 % глюкози витрачається |
|
|
|
не в гліколізі, а в пентозофосфатному шляху об- |
На частку фруктози припадає значна кіль- |
|
міну вуглеводів), а в м’язовій тканині — всього |
кість вуглеводів, що надходять з їжею (за добу — |
|
0,3 %. У печінці пентозофосфатний шлях стано- |
близько 100 г). Джерелами фруктози в їжі є фрук- |
|
вить 2,5–3 % від гліколізу, тобто приблизно в |
ти, мед і дисахарид сахароза, що у тонкому ки- |
|
10 разів більше, ніж у м’язах. |
шечнику перетворюється на глюкозу і фруктозу. |
|
3. В анаеробній фазі пентозофосфатного шля- |
Фруктоза після глюкози є найбільшим джерелом |
|
ху обміну вуглеводів утворюється фруктозо-6- |
вуглеводів, вона може включатися в гліколіз. |
|
фосфат, гліцеральдегід-3-фосфат і діоксіацетон- |
У жировій тканині фруктоза під дією гексокі- |
|
фосфат, які забезпечують взаємозв’язок гліколі- |
нази і за участю АТФ перетворюється на фрук- |
|
зу й пентозофосфатного шляху обміну вугле- |
тозо-6-фосфат, який під впливом фосфофруктокі- |
|
водів. |
|
|
|
|
|
|
|
|
|
|
|
нази і ще однієї молекули АТФ перетворюється |
|
4. В окисній фазі пентозофосфатного шляху |
на фруктозо-1,6-бісфосфат і далі по шляху гліко- |
|
(яка активна у присутності кисню) відбуваєть- |
лізу, або за участю гексозофосфатізомерази пере- |
|
ся синтез фосфопентоз і відновлених еквівалентів |
творюється на глюкозо-6-фосфат, що бере участь |
|
НАДФ+; у неокисній фазі (при зниженні вмісту |
у гліколізі, синтезі глікогену, пентозофосфатному |
|
кисню у тканині) утворення відновлених форм |
шляху або утворенні глюкози. |
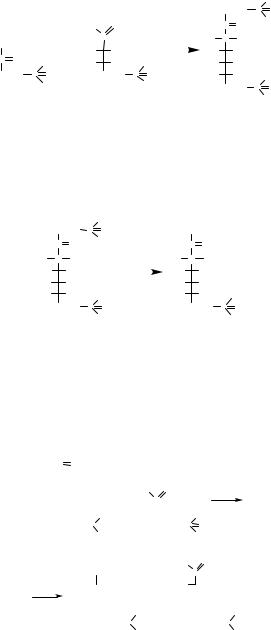


 Гліколіз, синтез глікогену, пентозофосфатний шлях
Гліколіз, синтез глікогену, пентозофосфатний шлях