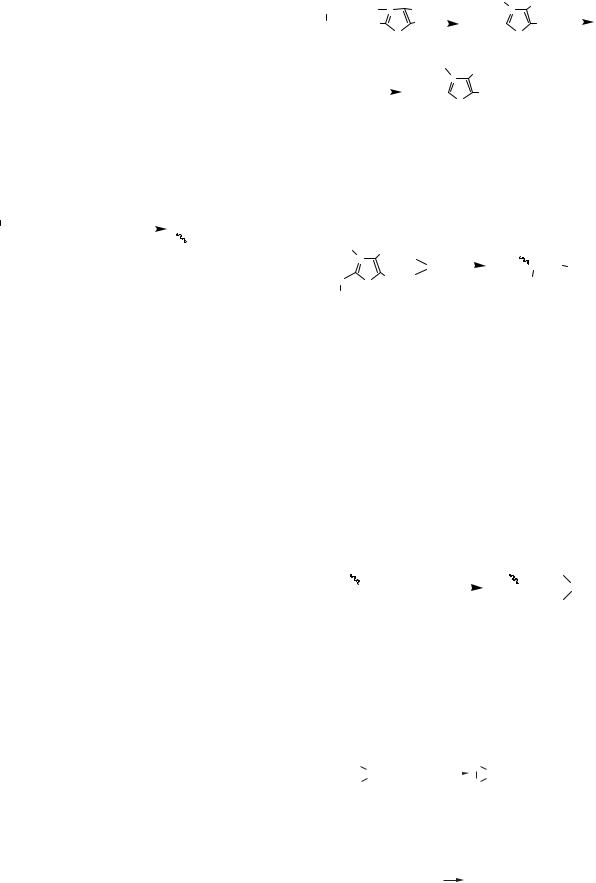|
ту, його називають ще анаеробним гліколізом. В |
Однак існують докази, що лактат, нагрома- |
|
аеробних умовах гліколіз являє собою лише пер- |
джений у м’язах під час роботи, може залучати- |
|
шу стадію повного аеробного розщеплення глю- |
ся до глюкогеногенезу, не залишаючи м’язів. |
|
кози до СО2 |
і Н2О. На другій стадії аеробного |
На етапі анаеробного гліколізу вивільняєть- |
|
розщеплення глюкози до СО2 |
і Н2О піруват |
ся 6–7 % енергії, що утворюються при повному |
|
піддається окисному декарбоксилюванню, тобто |
окисненні глюкози. На першій стадії гліколізу з |
|
втрачає СО2, а двовуглецевий фрагмент пере- |
однієї молекули глюкози утворюються дві моле- |
|
творюється на ацетил-КоА. Далі ця ацетильна |
кули гліцеральдегід-3-фосфату, окиснення якого |
|
група окиснюється до СО2 і Н2О в ЦТК. |
до пірувату сполучене з синтезом двох молекул |
|
Гліколіз і глікогеноліз перебігають у цито- |
АТФ: |
|
плазмі клітини, де локалізовані всі вищевказані |
а) у фосфогліцераткіназній реакції за рахунок |
|
ферменти цих процесів. Із 10 реакцій гліколізу три |
|
енергії 1,3-бісфосфогліцерату; |
|
реакції є необоротними: гексокіназна, фосфо- |
б) у піруваткіназній реакції за рахунок енергії |
|
фруктокіназна і піруваткіназна. |
|
|
|
|
|
|
|
|
|
|
|
фосфоенолпірувату. |
|
Сумарна реакція гліколізу: |
|
|
|
|
|
|
|
|
|
|
|
Отже, в анаеробному гліколізі утворюються |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
С6Н12О6 + 2АТФ + 2АДФ + 2Н3РО4 → |
4 молекули АТФ. Оскільки 2 молекули АТФ спо- |
|
живаються на першій стадії гліколізу (гексокіназ- |
|
Глюкоза |
|
|
|
|
|
|
|
|
|
на і фосфофруктокіназна реакції), енергетична |
|
|
→ 2С3Н6О3 + 4АТФ + 2Н2О |
роль анаеробного гліколізу полягає в синтезі |
|
|
2 молекул АТФ на 1 молекулу глюкози. |
|
|
|
|
|
Лактат |
|
|
|
|
|
1. Енергетична роль аеробного гліколізу така. |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Біологічна роль гліколізу. Енергетична функція |
Одним із продуктів гліколізу є НАДН+Н+, що |
|
утворюється при окисненні гліцеральдегід-3-фос- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Анаеробний гліколіз — один із процесів, що |
фату, причому на кожну молекулу глюкози ут- |
|
забезпечує клітини енергією в анаеробних умо- |
ворюються 2 молекули НАДН+Н+. В аеробних |
|
вах. Оскільки перші живі організми з’явилися на |
умовах НАДН+Н+ окиснюється за рахунок кис- |
|
Землі в той час, коли її атмосфера ще не містила |
ню, тобто атоми Гідрогену передаються не на |
|
кисню, анаеробний гліколіз слід вважати най- |
піруват, як це відбувається в анаеробному глі- |
|
давнішим із біологічних механізмів, призначених |
колізі, а на ланцюг дихальних ферментів. При |
|
для отримання енергії з органічних речовин. З |
окисненні однієї молекули НАДН+Н+ у ланцюзі |
|
усіх тканин анаеробний гліколіз найінтенсивні- |
дихальних ферментів синтезуються 3 молекули |
|
ше відбувається в м’язах під час роботи (наприк- |
АТФ, а при окисненні 2 молекул НАДН — 6 мо- |
|
лад, біг спортсмена на коротку дистанцію). Ки- |
лекул АТФ. Отже, на першому етапі енергетич- |
|
сень не встигає швидко надходити в м’язи і за- |
на ефективність аеробного гліколізу більша, ніж |
|
безпечувати окиснення пірувату і сполучений з |
анаеробного (вона становить 6 + 2 = 8 молекул |
|
ним синтез АТФ. У цих умовах м’язи використо- |
АТФ, якщо функціонує малатаспартатний чов- |
|
вують як «паливо» наявний запас глікогену і ге- |
никовий механізм, а якщо гліцерофосфатний, — |
|
нерують АТФ за допомогою анаеробного гліко- |
то 6 молекул АТФ). Враховуючи, що 2 молеку- |
|
лізу, кінцевим продуктом якого є лактат. Тому |
ли ацетил-КоА, утворені в аеробних умовах із |
|
під час бігу на коротку дистанцію в крові нагро- |
пірувату, окиснюючись у ЦТК, дають по 12 мо- |
|
маджується значна кількість лактату. Під час |
лекул АТФ кожна, то енергетична цінність |
|
відпочинку цей лактат перетворюється в печінці |
аеробного окиснення глюкози до СО2 і Н2О до- |
|
на глюкозу, яка надходить у м’язи (цикл Корі). |
рівнює 36–38 молекул АТФ. |
|
|
|
|
|
|
|
|
|
|
|
|
|
Енергетична роль глікогенолізу в перерахун- |
|
М’язи |
|
|
Кров |
|
|
|
Печінка |
|
ку на одну молекулу глюкози рівноцінна анае- |
|
|
|
|
|
|
|
|
|
|
|
|
|
робному або аеробному гліколізу. Під час гліко- |
|
Глюкоза |
|
|
|
|
|
|
Глюкоза |
|
|
|
|
|
|
|
|
|
генолізу АТФ не витрачається на утворення Гл- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
6-Ф, однак у процесі синтезу глікогену з глюко- |
|
Піруват |
|
|
|
|
|
|
Піруват |
|
|
|
|
|
|
|
|
|
зи одна молекула АТФ витрачається на утво- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
рення Гл-6-Ф із глюкози, що перетворюється на |
|
Лактат |
|
|
|
|
|
|
|
Лактат |
|
|
|
|
|
|
|
|
|
|
Гл-1-Ф, а останній, як відомо, використовується |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
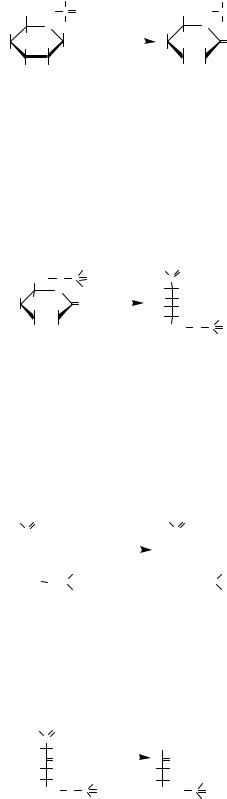
Крім лактату, з пірувату в м’язах може утво- |
на біосинтез глікогену. Крім того, 1 молекула |
|
УТФ витрачається на утворення УДФ-глюкози. |
|
рюватися шляхом трансамінування аланін, що |
2. Гліколіз готує «напівфабрикати» (піруват |
|
надходить у кров, печінку, де перетворюється на |
|
і лактат), які далі окиснюються в аеробних умо- |
|
піруват, а останній — на глюкозу (глюкозоала- |
вах ЦТК до СО2 і H2O з вивільненням значної |
|
ніновий цикл). |
|
|
|
|
|
кількості енергії. Як відомо, на кожну молекулу |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
глюкози в гліколізі утворюються 2 молекули |
|
М’язи |
|
|
Кров |
|
|
|
Печінка |
|
|
|
|
|
|
|
|
пірувату або 2 молекули лактату. При їхньому |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Глюкоза |
|
|
|
|
|
|
Глюкоза |
|
|
|
|
|
|
|
|
|
окисненні до СО2 і Н2О вивільнюється 93–94 % |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
від усієї енергії окиснення 1 молекули глюкози. |
|
Піруват |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Піруват |
|
2,3-Бісфосфогліцератний цикл. В еритроцитах |
|
|
|
|
|
|
|
|
|
|
|
|
|
багатьох ссавців є фермент бісфосфогліцератму- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Аланін |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
Аланін |
|
таза, який каталізує перетворення 1,3-бісфосфо- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|

 O
O