Материал: Біологічна та біоорганічна хімія_Мардашко О.О._ изд. 2008-342 с._ОНМедУ-2012
Утворення складних ефірів:
|
CH2OH |
|
|
|
|
|
|
|
||
|
|
O |
|
|
|
|
|
|
|
|
|
OH |
|
+ 5(CH3CO)2O |
|
|
|||||
|
|
|
||||||||
|
|
|
|
|
|
|
|
|||
OH |
|
|
OH |
|
|
|
|
|
|
|
|
|
OH |
|
|
|
|
|
|
|
|
|
|
O |
Ac |
|
|
|
|
|
|
|
|
|
CH2 |
|
|
|
|
|
|
||
|
|
|
|
|
O |
|||||
|
|
O |
Ac |
|
|
|
+ 5CH3COOH , |
|||
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|||
Ac O |
|
|
|
O |
|
Ac |
||||
|
|
|
|
|||||||
O Ac
де Ас — залишок оцтової кислоти.
Ефіри неорганічних кислот, зокрема ефіри фосфорної кислоти — фосфати — містяться у всіх рослинних і тваринних організмах, вони являють собою метаболічно активні форми моносахаридів. До них належать фосфати D-глюкози і D-фруктози. У печінці є фермент глюкокіназа, що каталізує фосфорилування глюкози (ефір фосфорної кислоти) по 6-му атому Карбону з утворенням глюкозо-6-фосфату. Ця ж сполука утворюється в м’язах за допомогою ферменту гексокі-
нази.
|
|
|
|
|
|
|
|
|
|
|
OH |
|
|
CH2OH |
|
|
CH2O |
|
P |
|
O |
||||
|
|
|
|
|
||||||||
|
|
|
|
|||||||||
|
|
O |
+АТФ |
|
O |
|
OH |
|||||
|
OH |
-АДФ |
|
|
|
|
|
|
|
|||
|
|
|
OH |
|
|
|
||||||
|
Глюкокіназа |
OH |
||||||||||
|
|
|
|
|
|
|
|
|||||
HO |
|
|
OH |
Гексокіназа HO |
|
|
|
|
||||
|
|
|
OH |
|||||||||
|
|
OH |
|
|
|
|
|
|
||||
D-Глюкоза D-Глюкозо-6-фосфат
Глюкозо-6-фосфат у повністю протонованій формі може існувати в середовищах із рН нижче 6. В організмі рН дорівнює 7,3, тому глюкозо-6- фосфат перебуває переважно у формі діаніону.
O-
CH2O P  O
O
O O-
OH
HO
OH
Фруктоза під дією ферменту гексокінази в організмі людини перетворюється на фруктозо-6- фосфат, а під дією ферменту печінки фруктокінази — переважно на фруктозо-1-фосфат.
D-Фруктозо-6-фосфат
O
HO P OH
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
а |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
з |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
а |
|
|
CH OH |
|
|
|
|
|
|
|
ін |
|
|
|||||
|
|
|
|
|
|
к |
|
|
|
||||||
|
2 |
|
|
|
|
|
|
|
о |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
с |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
к |
|
|
|
|
|
|
|
|
|
O |
|
|
|
е |
|
|
|
|
|
|
|
||
|
|
CH2OH Г |
|
|
|
|
|
|
|
|
|||||
|
|
HO |
|
Фр |
|
|
|
|
|
|
|
|
|
||
|
|
|
OH |
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
укт |
|
|
|
|
|
|
|||||
|
|
|
|
|
( |
|
|
|
о |
|
|
|
|
|
|
|
|
|
|
|
п |
|
|
|
кі |
|
|
|
|||
|
|
|
|
|
|
е |
|
|
|
н |
|
|
|||
|
OH |
|
|
|
|
|
|
а |
з |
|
|||||
|
|
|
|
чі |
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
н |
|
|
|
а |
||
D-Фруктоза |
|
|
|
|
|
|
ка) |
|
|
||||||
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
||
|
CH2O |
|
|
|||
|
O |
CH2OH |
||||
|
|
|
HO |
|
|
|
|
|
|
|
|
OH |
|
|
OH |
|||||
|
|
|
||||
CH OH |
|
|
|
|
OH |
|||||
|
2 O |
|
CH2O |
|
|
|
|
O |
||
|
|
|
P |
|
||||||
|
|
|
|
|||||||
|
|
|
|
|||||||
|
|
HO |
|
|
|
|
|
|
|
|
|
|
|
|
|
OH |
|||||
|
|
|
|
OH |
|
|||||
|
OH |
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|||
D-Фруктозо-1-фосфат
Відновлення
H |
|
C |
|
|
O |
|
|
|
|
CH2OH |
||||||
|
|
|
|
|
|
|||||||||||
|
|
|
|
|
|
|
|
H |
|
|
|
|
OH |
|||
H |
|
|
C |
|
|
OH Aмальгама натрію |
|
C |
|
|
||||||
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
||||
HO |
|
|
C |
|
|
H |
H2/Ni, LiAIH4 |
HO |
|
C |
|
|
H |
|||
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
H |
|
|
C |
|
|
OH |
|
H |
|
C |
|
OH |
||||
|
|
|
|
|
|
|
||||||||||
|
|
|
|
|
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
H |
|
|
C |
|
OH |
|
H |
|
C |
|
OH |
|||||
|
|
|
|
|
|
|||||||||||
|
|
|
|
|||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
CH2OH |
|
|
|
CH2OH |
||||||||
D-Глюкоза D-Сорбіт
При відновленні цукрів утворюються багатоатомні спирти. Цукрові спирти — це безбарвні солодкі речовини, які легко розчиняються у воді.
Сорбіт є вихідним продуктом для синтезу аскорбінової кислоти (вітамін С). Він замінює цукор хворим на діабет.
Окиснення Залежно від природи окисника та умов ре-
акції (рН середовища, температури) моносахариди перетворюються на різні продукти. Продуктами окиснення цукрів є полігідроксикислоти (альдонові за дії м’яких окисників і глікарові, якщо окисником є HNO3 чи KMnO4).
H |
O |
HO |
O |
HO |
O |
|||||||||||||||||||||
|
|
|
C |
|
|
|
|
|
|
|
C |
|
|
|
|
|
|
|
C |
|
|
|
|
|||
H |
|
|
|
|
|
|
OH |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
C |
|
|
H |
|
|
C |
|
|
OH |
H |
|
|
C |
|
|
OH |
||||||||
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
|
|
|
|
|
|
|
H |
Br2 + H2O(HBrO) |
|
|
|
|
|
|
|
H HNO3[O] |
|
|
|
|
|
|
|
|
|
|
HO |
|
C |
|
|
HO |
|
C |
|
|
HO |
|
C |
|
|
|
H |
||||||||||
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
H |
|
C |
|
OH |
H |
|
C |
|
OH |
H |
|
C |
|
|
|
OH |
||||||||||
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
H |
|
C |
|
OH |
H |
|
C |
|
OH |
H |
|
C |
|
|
|
OH |
||||||||||
|
|
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
CH2OH |
|
|
|
|
CH2OH |
HO |
|
C |
|
|
|
O |
|||||||||||
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
І |
ІІ |
ІІІ |
|
Глюконова |
Глюкарова |
D-глюкоза |
(альдонова) |
(цукрова) |
|
кислота |
кислота |
Ці реакції відбуваються в кислому чи нейтральному середовищі.
Якщо реакція перебігає в лужному середовищі, то утворюється суміш продуктів окиснення вуглеводів. Ці реакції застосовуються для якісного визначення положення карбонільної групи у вуглеводах. Навіть такі слабкі окисники, як гідроксид срібла в аміачному середовищі (реактив Толленса) чи лужний розчин тартратного комплексу Купруму (ІІ) (реактив Фелінга), легко відновлюються альдозами.
Реакція срібного дзеркала:
H |
|
|
t° |
|
|
суміш продуктів |
|
O t°, [Ag(NH ) ]OH |
|
+ |
окиснення |
||
|
C |
|
3 2 |
Ag |
|
|
R
Альдоза
Реакція Фелінга:
|
|
COOK |
t° |
O |
|
|
|
COOK |
|||
O |
|
|
CHO |
|
t0, 2H O |
|
|
|
|
CHOH |
|
|
|
|
|
|
|
|
|||||
R C + 2 |
|
|
Cu |
2 |
R C |
+ Cu2O |
|
+ 2 |
|
|
|
CHO |
|
|
CHOH |
||||||||
H |
|
|
|
OH |
|||||||
|
|
COONa |
|
|
COONa |
||||||
|
|
|
|
|
|
|
|
|
|||
Альдоза |
Реактив |
|
Червоний |
|
|
||||||
Фелінга |
|
|
осад |
|
|
||||||
Суміш продуктів окиснення
59
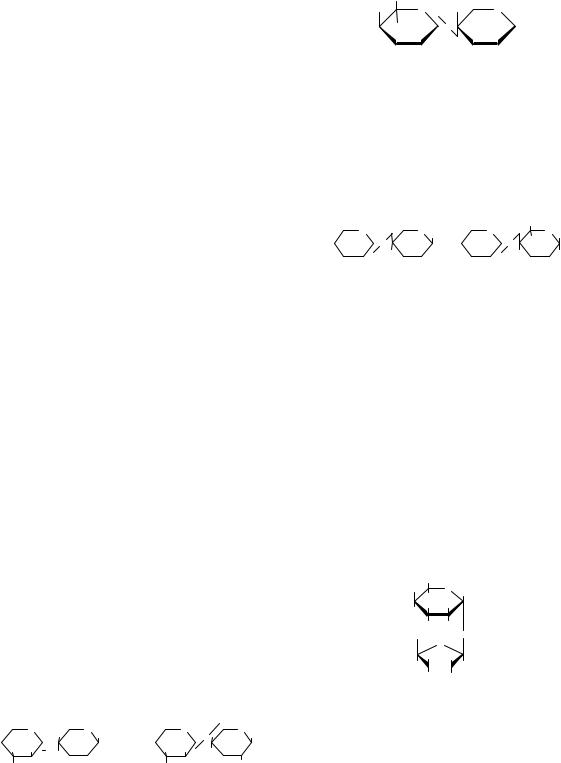
3.2. СТРУКТУРА І ФУНКЦІЇ |
|
|
|
|
|
|
|
|
|
Целобіоза |
|
|
|
|
|
|
|
|
|||||||||||||
ОЛІГО- І ПОЛІСАХАРИДІВ |
Целобіоза є проміжним продуктом гідролізу |
||||||||||||||||||||||||||||||
|
|
|
|
|
|||||||||||||||||||||||||||
Дисахариди |
|
|
|
|
клітковини. При гідролізі утворює дві молекули |
||||||||||||||||||||||||||
|
|
|
|
глюкози. Це відновлювальний цукор, може бути |
|||||||||||||||||||||||||||
Дисахариди (олігосахариди) складаються з |
окисненим у целобіонову кислоту. Відмінність від |
||||||||||||||||||||||||||||||
двох моносахаридних ланок однакової або різної |
мальтози полягає в тому, що аномерний атом, |
||||||||||||||||||||||||||||||
природи. Легко гідролізуються. Загальна форму- |
який бере участь в утворенні глікозидного зв’яз- |
||||||||||||||||||||||||||||||
ла дисахаридів така: С |
12 |
Н |
22 |
О . Це кристалічні |
ку, має β -конфігурацію. |
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
|
|
11 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
речовини. При розчиненні у воді вони утворю- |
|
|
|
|
|
|
|
CH2OH |
|
|
|
|
|
CH2OH |
|
|
|
|
|
||||||||||||
ють істинні розчини, солодкі на смак. Основні біо- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
|
|
|
|
|
|
|
|
O |
|
|
|
|
|
|
|
|
O |
OH |
|
|
|
|||||||||
зи, при гідролізі яких утворюються монози, на- |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O |
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
OH |
|
|
|
|
OH |
|
|
|
|
|
|
|
|
|||||||||||
ведені нижче: |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
HO |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
Целобіоза + Н2О → D-глюкоза + D-глюкоза |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||
|
|
|
|
|
|
|
|
|
OH |
|
|
|
OH |
|
|
|
|||||||||||||||
Мальтоза + Н2О → D-глюкоза + D-глюкоза |
β -D-Глюкопіранозил-1,4-β -D-глюкопіраноза |
||||||||||||||||||||||||||||||
Лактоза + Н2О → D-глюкоза + D-галактоза |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
Сахароза + Н2О → D-глюкоза + D-фруктоза |
|
|
|
|
Лактоза (молочний цукор) |
||||||||||||||||||||||||||
Дисахариди належать до О-глікозидів, у яких |
Лактоза (β -D-галактопіранозил-1,4-β -D-глю- |
||||||||||||||||||||||||||||||
друга молекула моносахариду відіграє роль аг- |
копіраноза) міститься в молоці (4–5 %). Вона |
||||||||||||||||||||||||||||||
лікону. За типом зв’язування моносахаридних |
складається із залишків β -D-галактопіранози і |
||||||||||||||||||||||||||||||
залишків дисахариди можна розподілити на дві |
D-глюкози, які з’єднані β -зв’язком. |
|
|
|
|||||||||||||||||||||||||||
групи: |
|
|
|
|
Галактоза |
Глюкоза |
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
1. Зв’язок утворюється за рахунок напіваце- |
|
CH2OH |
|
|
|
|
|
|
|
|
|
CH2OH |
|
|
|
|
|
||||||||||||||
тальної (глікозидної) ОН-групи одного і будь- |
OH |
|
O |
|
|
O |
|
|
|
OH |
|
|
O |
|
|
|
O OH |
||||||||||||||
якої спиртової ОН-групи іншого моносахариду |
|
OH |
|
|
|
O |
|
OH |
|
|
|
|
|
|
OH |
|
|
|
O |
|
OH |
||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
OH |
|
|
|
|
|
|
|
|
|
|
|
||||||||
(відновлювальні дисахариди). |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||||
|
|
|
OH |
|
OH |
|
|
OH |
|
|
|
OH |
|||||||||||||||||||
2. Зв’язок утворюється за рахунок напіваце- |
|
|
α |
-Лактоза |
|
|
|
|
|
|
|
β -Лактоза |
|||||||||||||||||||
тальних (глікозидних) ОН-груп обох моносаха- |
|
|
|
|
|
|
|
|
|
||||||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
ридів (невідновлювальні дисахариди). |
Лактоза застосовується в фармацевтичній |
||||||||||||||||||||||||||||||
|
|
|
|
|
|||||||||||||||||||||||||||
Відновлювальні дисахариди |
промисловості для виготовлення порошків і таб- |
||||||||||||||||||||||||||||||
У відновлювальних дисахаридів в утворенні |
леток, вона є складовою частиною продуктів |
||||||||||||||||||||||||||||||
глікозидного зв’язку бере участь напівацеталь- |
харчування для малюків. У материнському мо- |
||||||||||||||||||||||||||||||
ний гідроксил однієї монози і спиртовий гідроксил |
лоці вміст лактози сягає 8 % від маси. |
||||||||||||||||||||||||||||||
другої, найчастіше у С-4 або С-6, іноді у С-3. При |
|
|
Невідновлювальні дисахариди |
||||||||||||||||||||||||||||
цьому одна напівацетальна ОН-група зали- |
|
|
|||||||||||||||||||||||||||||
Представником цього ряду дисахаридів є са- |
|||||||||||||||||||||||||||||||
шається вільною і зберігає здатність до розкрит- |
|||||||||||||||||||||||||||||||
тя циклу (цикло-оксо-таутомерія). Свіжоприго- |
хароза (буряковий і тростинний цукор). До її |
||||||||||||||||||||||||||||||
товлені розчини таких дисахаридів мутаротують |
складу входять α -D-глюкоза і β -D-фруктоза: |
||||||||||||||||||||||||||||||
і реагують із реактивами на альдегідну групу |
глюкоза — у піранозній, а фруктоза — у фура- |
||||||||||||||||||||||||||||||
аналогічно глюкозі: відновлюються в багато- |
нозній формі. Глікозидний зв’язок утворюється за |
||||||||||||||||||||||||||||||
атомні спирти, окиснюються в альдонові кисло- |
рахунок гідроксильних груп при аномерних ато- |
||||||||||||||||||||||||||||||
ти. Для відновлювальних цукрів характерною є |
мах Карбону. Тому сахароза не здатна до цик- |
||||||||||||||||||||||||||||||
цикло-оксо-таутомерія. |
|
|
|
|
ло-оксо-таутомерії і не виявляє відновних влас- |
||||||||||||||||||||||||||
Мальтоза (солодовий цукор) |
тивостей (не реагує з реактивом Толленса і реак- |
||||||||||||||||||||||||||||||
тивом Фелінга). |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||||
Дисахарид мальтоза (α -D-глюкопіранозил- 1,4-α -D-глюкопіраноза) складається із залишків двох α -глюкоз, зв’язок α -1,4-глікозидний. У назві цієї сполуки перша молекула має закінчення «озил», якщо в реакції брав участь напівацетальний гідроксил, а друга молекула зберігає закінчення «оза».
Утворюється як проміжний продукт при дії амілаз на крохмаль.
|
|
CH2OH |
|
|
|
|
CH2OH |
|
|
CH2OH |
|
|
CH |
|||||||||||
|
|
O |
|
|
|
|
O |
|
|
O |
|
O |
|
2 O |
||||||||||
|
OH |
|
|
|
O |
|
OH |
|
|
|
|
OH |
|
|
|
OH |
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
OH |
|
|
|
OH |
OH |
|
OH |
|
OH |
|||||||||||||||
|
|
|
|
|
|
|
|
|||||||||||||||||
|
|
OH |
|
|
|
|
OH |
|
|
OH |
|
|
|
OH |
||||||||||
|
|
Мальтоза |
|
|
Ізомальтоза |
|||||||||||||||||||
α -1,4-Глікозидний зв’язок |
α -1,6-Глікозидний зв’язок |
|||||||||||||||||||||||
CH2OHO
HO OH
OH CH2OHO O
OH
 CH2OH OH
CH2OH OH
α -D-Глюкопіранозил-1,2-β -D- фруктофуранозид (сахароза)
При гідролізі сахарози, яка має праве обертання, знак обертання змінюється, тому що утворюється D-фруктоза, яка значно сильніше обертає вліво, ніж D-глюкоза — вправо. Це явище називається інверсією, а одержана суміш —
інвертним цукром (штучний мед).
60
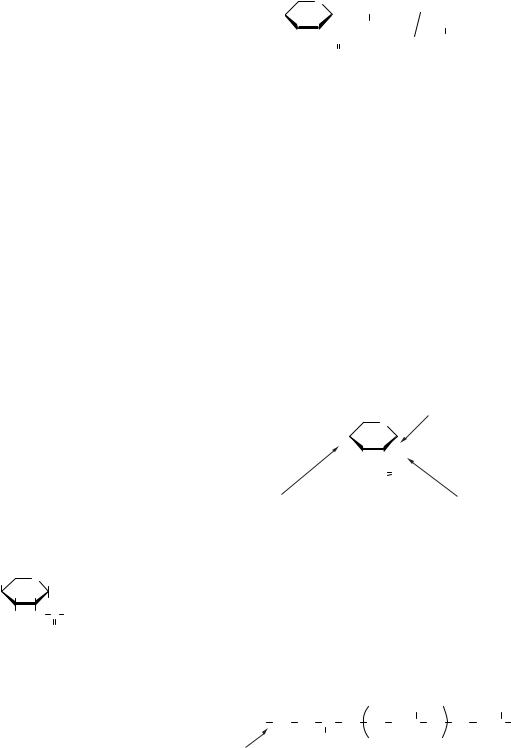
Ферменти людського організму здатні розщеплювати 1,4- і 1,6-глікозидні зв’язки в крохмалі та глікогені. Відмічено, що гідроліз сахарози, на відміну від інших біоз, перебігає легше, оскільки фруктоза, що входить до її складу, перебуває у вигляді менш стійкого п’ятичленного фуранозного циклу. Сахароза легко гідролізується в кишечнику і, подібно до моносахаридів, потрапляє у кров. Гідроліз іншого дисахариду — лактози
— відбувається сповільнено внаслідок зниження активності ферменту, що розщеплює лактозу, — лактази. Зниження активності ферменту призводить до непереносності деякими людьми молока та молочних продуктів харчування. Олігосахариди досить поширені в природі. У рослинному світі вони відіграють роль резервних вуглеводів. Найчастіше зустрічаються олігосахариди групи сахарози. Майже так широко, як сахароза, у рослинах поширені рафіноза і стахіоза.
Другою великою групою природних олігосахаридів є олігосахариди молока, яким належить важлива роль у формуванні кишкової флори новонароджених, необхідної для травлення. Вони сприяють розвитку в шлунково-кишковому тракті мікроорганізму Lactobacillus bifidus, який розщепляє лактозу молока (дисахарид) з утворенням молочної й оцтової кислоти, яка перешкоджає росту патогенних бактерій. До їхнього складу входять D-галактоза, α -фруктоза, N-ацетилглюкозамін і залишок лактози. Один із важливих олігосахаридів молока — лакто-N-фу- копентоза.
Глікопротеїни
Глікопротеїни — це змішані біополімери, в яких молекула білка містить ковалентно приєднані олігосахаридні ланцюги.
CH2OH |
|
|
O |
|
|
|
NH2 |
|||||
|
O NH |
|
|
|
|
|
|
|
|
|
|
|
|
|
C |
|
CH |
2 |
CH |
|
COOH |
||||
|
|
|
|
|
||||||||
|
|
|
OH |
|
OH |
|
|
Аспарагін |
|
NH C CH3
O
N-Глікопротеїн
Глікопротеїни входять до складу всіх органів, тканин і клітин організму людини і тварин; вони містяться в секреторних рідинах і плазмі крові. Функції глікопротеїнів надзвичайно різноманітні. Серед них зустрічаються ферменти, гормони, білки імунної системи, компоненти плазми крові, муцини, рецептори клітинних мембран. Найбільш розповсюдженим типом зв’язку у тваринних глікопротеїнах є N-глікозидний зв’язок, утворений залишками N-ацетилглюкозаміну і β -амідною групою аспарагіну (імуноглобуліни, антигени тканинної сумісності).
Іншим типом зв’язку у тваринних глікопротеїнах є О-глікозидний зв’язок між залишками N-ацетилгалактозаміну і гідроксильною групою серину чи треоніну. Цей тип зв’язку характерний для муцинів, речовин груп крові, глікофорину мембрани еритроцитів і т. ін.
|
|
|
CH2OH |
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
OH |
|
|
O |
|
|
NH2 |
|
|
|
|
|
NH2 |
|||||||||||||
OH |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
|
O |
|
CH2 |
|
|
CH |
|
COOH |
HO |
|
CH |
|
CH |
|
COOH |
||||||
|
|
|
|
|
|||||||||||||||||||||
|
|
|
|
|
NH |
|
C |
|
CH |
3 |
|
|
|
|
|
|
CH3 |
||||||||
|
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
O
О-Глікопротеїн
У багатьох глікопротеїнах (як сироваткових, так і мембранних) одночасно присутні О- і N-глікозидні ланцюги.
Синтез вуглеводної частини глікопротеїнів
1. О-зв’язані глікопротеїни. Поліпептидні лан-
цюги О-зв’язаних глікопротеїнів кодуються відповідними мРНК; оскільки більшість глікопротеїнів зв’язана з мембранами, вони звичайно транслюються на мембранозв’язаних полірибосомах. Олігосахаридні ланцюги глікопротеїнів О-глікозидного типу конструюються шляхом
східчастого додавання цукрів із нуклеотидцукрів,
таких як УДФ-GalNAc, УДФ-Gal, ЦДФ NeuAc.
Ферменти, які каталізують цю реакцію, являють собою мембранозв’язану глікопротеїн-глікозил- трансферазу. Для утворення одного типу зв’язку потрібна відповідна трансфераза (гіпотеза: «один зв’язок — одна глікозилтрансфераза»).
|
|
CH2OH |
|
|
Серин |
||||
OH |
|
O |
|
|
|
|
|
||
|
OH |
|
|
|
~ |
||||
|
|
|
|
|
|
||||
|
|
|
|
O |
|
CH2 |
|
C |
|
|
|
|
NH |
~ |
|||||
|
|
|
|
|
|
||||
|
|
|
C O |
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
|
|
|
CH3 |
|
|
|
|
||
N-Ацетилгалактозамін |
|
|
|
|
О-Глікозидний зв’язок |
||||
О-зв’язані глікопротеїни
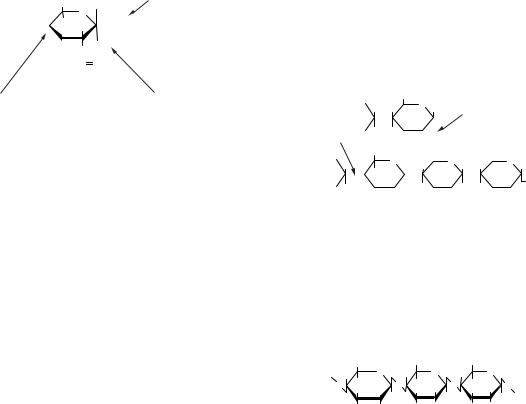
2. N-зв’язані глікопротеїни. Синтез полісаха-
ридів, приєднаних N-глікозидним зв’язком, відбувається інакше, у цьому випадку бере участь проміжний носій олігосахариду доліхолфосфат. Це амфіфільна речовина. Гідрофобним кінцем вона занурена в мембрану, а виступаючий гідрофільний кінець служить акцептором першого моносахариду — GlcNAc; у результаті реакції утворюється N-ацетилглюкозаміндифос- фатдоліхол.
CH3 |
CH3 |
HO CH2 CH2 CH CH2 CH2 CH C CH2 CH2 CH
C CH2 CH2 CH C CH3
C CH3
CH3 |
n |
|
|
Приєднання фосфату |
Доліхол n=17–20 |
Потім при дії серії специфічних глікозилтрансфераз приєднуються інші моносахари, утворюється олігосахарид. Він переноситься на амідну групу аспарагіну в складі пептидного ланцюга (глікопротеїну) в процесі їх утворення на мембранних полірибосомах. Далі під дією окремих глікозилтрансфераз, локалізованих головним чином в апараті Гольджі, відбувається приєднання вуглеводного фрагмента.
61
|
|
CH2OH |
|
|
|
Аспарагін |
|||||
|
|
|
|
|
|
|
|
|
|||
HO |
|
O |
|
|
|
|
|
|
|
||
|
|
|
|
|
|
|
|
|
|
||
|
|
OH |
|
O |
~ |
||||||
|
|
|
|
|
|
|
|
|
|
||
|
|
|
|
NH NH |
|
C |
|
CH2 |
|
C |
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
~ |
|
|
|
|
C O |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
CH3 |
|
|
|
|
|
|
|
|
N-Ацетилгалактозамін |
|
|
|
N-Глікозидний зв’язок |
|||||||
N-Зв’язані глікопротеїни
Найбільш поширені гліколіпіди — це глікоцераміди (глікосфінголіпіди), що являють собою похідні цераміду. Антигени А і В мембран еритроцитів є глікоцерамідами. Гліколіпіди зустрічаються тільки в мембранах (головним чином, у плазматичній мембрані).
Полісахариди
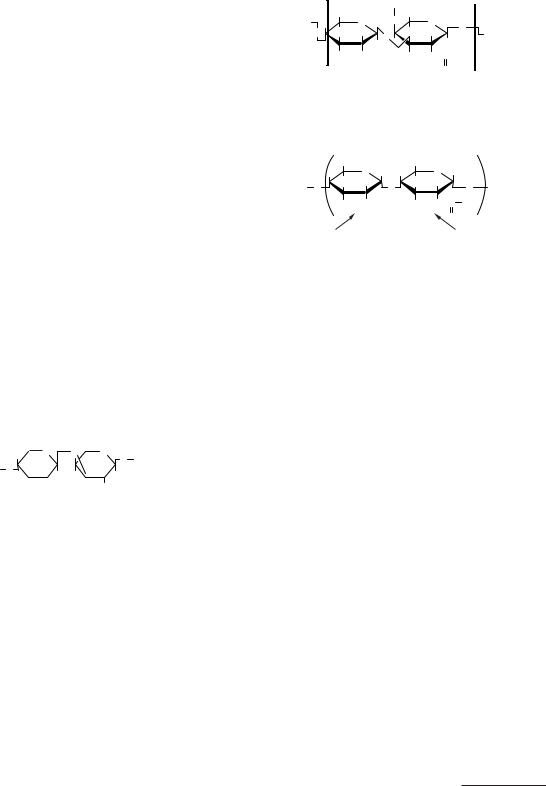
Полісахариди утворюють основну масу органічної речовини нашої планети. Досить згадати, що з них складається основна частина рослинного світу, де полісахариди виконують скелетні функції, а також служать резервними вуглеводами.
Гомополісахариди
Із вищих полісахаридів найбільш розповсюдженими і важливими є крохмаль, глікоген і клітковина. Усі три полісахариди мають загальну формулу (С6Н12О5)n і складаються із залишків глюкози. Однак їхні структури істотно розрізняються.
Крохмаль є запасним поживним вуглеводом рослин — тимчасове депо глюкози рослин. Складається з двох фракцій: амілози й амілопектину (табл. 3.1). Амілоза в крохмалі становить близько 20 %, амілопектин — близько 80 %.
|
Таблиця 3.1 |
Склад крохмалю |
|
|
|
Амілоза |
Амілопектин |
|
|
α -1,4-глікозидні зв’язки |
Крім α -1,4-глікозидних |
|
зв’язків є і α -1,6-гліко- |
|
зидні зв’язки, що забезпе- |
|
чують його форму. На 30 |
|
α -1,4 зв’язків припадає |
|
приблизно одинα -1,6- |
|
зв’язок |
Входить до складу 200– |
Входить до складу 600– |
1000 залишків глюкози |
6000 залишків глюкози |
Добре розчинна в гаря- |
Погано розчинний |
чій воді |
у гарячій воді |
Йодом забарвлюється в |
Йодом забарвлюється |
синій колір |
у фіолетовий колір |
Глікоген — основне депо вуглеводів (глюкози) в організмі тварин і людини. Глюкоза глікогену використовується організмом в інтервалах між прийманням їжі й у період м’язової активності. Особливо багаті на глікоген печінка і скелетні м’язи, де зосереджено близько 90 % всієї його кількості в організмі. У печінці глікогену міститься 2–5 % від загальної ваги органа (близько 150 г), у скелетних м’язах — 0,5–2 % (близько 200 г). Глікоген, як і крохмаль, складається із залишків α -глюкоз, з’єднаних α -1,4-глікозидними зв’язками,
але крім цього наявна значна кількість α -1,6-гліко- зидних зв’язків, що забезпечують гіллясту будову глікогену, тобто за будовою глікоген близький до амілопектину. Приблизно на кожен десяток молекул глюкози з α -1,4-глікозидними зв’язками припадає один α -1,6-глікозидний зв’язок (рис. 3.3).
|
|
|
|
|
|
|
|
6 CH2OH |
|
α |
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
1 |
|
4 |
|
5 |
O |
|
(1,6)-глікозидні |
||||||||||||
|
|
|
|
|
|
|
||||||||||||||||||
|
|
|
|
|
|
|
|
|
|
|||||||||||||||
α (1,4)-глікозидні |
O |
OH |
|
1 |
|
|
|
|
|
зв’язки |
||||||||||||||
|
|
|
3 |
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
2 O |
|
|
|
|
|
|
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
||||||
зв’язки |
|
|
|
|
|
|
OH |
|
|
|
|
|
|
|
|
|
|
|
|
|||||
6 |
CH2OH |
|
|
6 |
CH2 |
|
|
|
|
|
6 |
CH2OH |
||||||||||||
|
|
|
|
|
|
|
|
|
|
|
||||||||||||||
|
|
|
|
|
5 |
O |
|
|
|
|
5 |
O |
|
|
|
|
5 |
O |
||||||
|
1 4 |
OH |
|
|
1 |
4 OH |
|
|
1 4 |
OH |
1 |
|
|
|||||||||||
|
|
O |
|
3 |
|
|
|
2 |
|
O |
|
3 |
|
|
2 |
|
O |
|
3 |
|
|
2 O |
|
|
|
|
|
|
|
|
|
OH |
|
|
|
OH |
|
|
|
|
|
OH |
|||||||
Рис. 3.3. Будова крохмалю і глікогену
Целюлоза (клітковина) — лінійний полісаха-
рид, побудований з молекул β -глюкози, з’єднаних β -(1→ 4)-глікозидними зв’язками, ступінь полімеризації 2000–3000.
Звичайне її джерело — деревина, містить близько 50 % целюлози.
|
CH OH |
|
CH2OH |
|
CH2OH |
|
|
|
2 |
O |
|
O |
|
O |
|
|
|
|
|
|
|||
O |
OH |
|
O |
OH |
O |
OH |
O |
|
|
|
|
|
|||
|
|
OH |
|
OH |
|
OH |
|
Целюлоза
Геміцелюлоза — суміш полісахаридів клітинної стінки рослин, склад яких залежить від виду рослини. Залежно від моносахаридного складу основного ланцюга геміцелюлози поділяються на ксилани, глюкоманани і галактани.
У наземних рослинах і водоростях широко представлені пектинові речовини — поліуроніди. Основним компонентом рослин є D-галактуроно- ва кислота, а в малих кількостях присутні L-ара- біназа і D-галактоза.
Розчинні пектинові речовини знаходяться, головним чином, у соках рослин, нерозчинні утворюють міжклітинну речовину і значну частину стінки молодих рослин. Частково етерифіковані поліуроніди називаються пектиновими кислота-
ми, а самі поліуроніди — пектовими кислотами.
Пектинові кислоти здатні утворювати в розчинах міцні гелітаколоїди, що зумовлено міжмолекулярною асоціацією. Це знаходить застосування в кондитерській і фармацевтичній промисловості.
Гетерополісахариди
Камедями називаються полісахариди, що виділяються при ушкодженні кори рослин у вигляді в’язких розчинів і перетворюються на склоподібну масу. Слизи — споріднені камедям полісахариди, присутні у неушкоджених рослинах. Джерелами слизів служать кора, листя, корені тощо. Слизи є продуктом метаболізму рослин. Камеді утворюються в результаті патологічних процесів
— механічного і бактеріального походження. Обидва типи утворень є гетерополісахаридами складного складу і будови.
62
Протеоглікани — сполуки, що складаються з білків і полісахаридів, у яких на частку полісахариду припадає основна частина молекули — звичайно більше 95 %. Вуглеводна частина в протеогліканах представлена глікозаміногліканами (кислими мукополісахаридами).
Глікозаміноглікани утворюються з повторюваних дисахаридних залишків, кожний з яких є похідною аміногексози — звичайноD-глюкозамінучи D-галактозаміну. Щонайменше один із двох цукрів у повторюваному дисахаридному залишку глікозаміноглікану містить кислотну групу, що має при рН=7 негативний заряд — як правило, це карбоксильна чи сульфатна група.
Протеоглікани присутні в гелеподібній основній речовині, що заповнює в більшості тканин простір між клітинами. Крім того, вони містяться в хрящах, сухожиллях і шкірі, а також у синовіальній рідині, що виконує функцію змащування в суглобах. Молекули протеогліканів у розчині «розпущені» внаслідок відштовхування одноіменно заряджених ланцюгів глікозаміногліканів (займають великий об’єм). При збільшенні тиску об’єм, який займають молекули, зменшується, тому що рідина вичавлюється з проміжків. Якщо тиск зняти, молекула знову набуває розпущеної форми. Ця властивість дуже важлива для суглобів.
Гіалуронова кислота — це глікозаміноглікан міжклітинної основної речовини тканин тварин, що складається із залишків D-глюкуронової кислоти і N-ацетил-D-глюкозаміну, які багаторазово чергуються.
|
|
COOH |
|
|
CH2OH |
|
|
|
|
|
||
|
|
HO |
O |
|
O |
|
O |
|||||
O |
OH |
HO |
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|||||
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
|
OH |
|
OH NH |
|
C |
|
CH3 |
||||
|
|
|
|
|
||||||||
|
|
|
|
|
|
|
||||||
|
|
|
|
|
|
|
|
|
O |
|||
Гіалуронова кислота
Гіалуронова кислота утворює дуже в’язкі, гелеподібні розчини. Протеоглікановий гель обмежує дифузію і проникність для молекул і частинок. Фермент гіалуронідаза, який секретується деякими патогенними бактеріями, здатний гідролізувати глікозидні зв’язки гіалуронової кислоти і тим самим полегшувати проникнення бактерій у тканину.
Хондроїтин-4-сульфат — основний компо-
нент протеогліканів хряща, що складається з
D-глюкуронової кислоти і N-ацетил-D-галактоз- 4-сульфату.
|
|
|
S3OH |
|
|
|||
COOH |
O |
CH2OH |
|
|
||||
|
O |
|
O |
|
O |
|||
|
|
|
|
|||||
O OH |
O |
|
|
|
|
|||
|
|
|
|
|
||||
|
|
|
|
|
|
|
|
|
|
OH |
|
|
NHC |
|
CH3 |
||
|
|
|
|
|||||
O
Хондроїтин-4-сульфат
Гепарин відомий як коагулянт, це — глікозаміноглікан. Він синтезується тучними клітинами.
|
COOH |
CH |
OSO |
H |
|||||
|
|
|
O |
2 |
O 3 |
|
|||
O |
OH |
|
O |
OH |
|
|
O |
||
|
|
|
|
|
|
|
|||
|
|
|
OSO3H |
|
|
NHC CH3 |
|||
|
|
|
|
|
|
|
|
O |
|
Залишок D-глюк- |
|
|
Залишок N-ацетил- |
||||||
уронат2-сульфату |
|
глюкозамін-6-сульфату |
|||||||
Гепарин
КОНТРОЛЬНІ ПИТАННЯ
1.Вуглеводи: визначення, класифікація. Моносахариди (альдози і кетози; тріози, тетрози, пентози, гексози, гептози); біомедичне значення окремих представників.
2.Моносахариди: пентози (рибоза, 2-дезок- сирибоза, ксилоза), гексози (глюкоза, галактоза, маноза, фруктоза) — будова, властивості. Якісні реакції на глюкозу.
3.Будова та властивості похідних моносахаридів. Амінопохідні: глюкозамін, галактозамін. Уронові кислоти. L-Аскорбінова кислота (вітамін С). Продукти відновлення моносахаридів: сорбіт, маніт.
4.Олігосахариди: будова, властивості. Дисахариди (сахароза, лактоза, мальтоза); їх біомедичне значення.
5.Полісахариди. Гомополісахариди: крохмаль, глікоген, целюлоза, декстрини — будова, гідроліз, біомедичне значення. Якісна реакція на крохмаль.
6.Гетерополісахариди: визначення, структура. Будова та біомедичне значення глікозаміногліканів (мукополісахаридів) — гіалуронової кислоти, хондроїтинсульфатів, гепарину.
Глава 4. БІОЛОГІЧНО АКТИВНІ ГЕТЕРОЦИКЛІЧНІ СПОЛУКИ.
НУКЛЕОЗИДИ, НУКЛЕОТИДИ, НУКЛЕЇНОВІ КИСЛОТИ
4.1. КЛАСИФІКАЦІЯ, БУДОВА ТА ЗНАЧЕННЯ БІОЛОГІЧНО ВАЖЛИВИХ ГЕТЕРОЦИКЛІЧНИХ СПОЛУК
Циклічні сполуки, які у складі циклу, крім атомів Карбону, містять інші атоми, дістали на-
зву гетероциклічних (від грецьк. heteros — інший). Найбільше значення мають гетероциклічні сполуки з атомами Нітрогену, Сульфуру й Оксигену. Валентні кути між зв’язками у цих атомів майже не відрізняються від валентних кутів атомів Карбону у стані sp3- або sp2-гібри-
63