Материал: 1010-pochvovedenie-2013-7

846 ДЕМКИНА и др.
Таблица 5. Влияние степени солонцеватости на изменения величин микробиологических параметров в различ& ных горизонтах подкурганных ( 2000 лет назад) почв
Горизонт |
Параметр |
Вклад в дисперсию, % |
Уровень значимости |
Группирование величин параметров по кри& |
|
|
|
F&распределения |
терию Дункана (α = 5%) |
А1 |
НА |
65.0 |
0.0314* |
А – каштановая несолонцеватая, каштано& |
|
|
|
|
вая сильносолонцеватая, |
|
|
|
|
АВ – каштановая солонцеватая, |
|
|
|
|
В – солонец |
|
БТМ |
67.6 |
0.0232* |
А – каштановая солонцеватая, |
|
|
|
|
АВ – каштановая сильносолонцеватая, |
|
|
|
|
В – солонец, каштановая несолонцеватая |
В1 |
БС |
86.0 |
0.0009*** |
А – каштановая солонцеватая, |
|
|
|
|
В – каштановая несолонцеватая, каштано& |
|
|
|
|
вая сильносолонцеватая, |
|
|
|
|
С – солонец |
|
БТМ |
80.6 |
0.0032** |
А – каштановая солонцеватая, |
|
|
|
|
В – каштановая несолонцеватая, каштано& |
|
|
|
|
вая сильносолонцеватая, солонец |
В2 |
ПА |
80.2 |
0.0035** |
А – каштановая солонцеватая, |
|
|
|
|
В – солонец, |
ВС – каштановая несолонцеватая, С – каштановая сильносолонцеватая
Таблица 6. Эколого&трофическая структура микробных сообществ современных и подкурганных почв солонцо& вых комплексов (по средневзвешенным в профиле величинам)
|
|
Σ численность |
ЭТС, % |
|
Индекс |
|
Время |
Почва |
микроорганизмов, |
БС/НА |
олиготрофности, |
||
(ПА : НА : БС) |
||||||
|
|
млн КОЕ/г почвы |
|
|
(ПА : БС × 100) |
|
Современность |
Каштановая солонцеватая |
13.36 |
54 : 7 : 39 |
5.7 |
139 |
|
|
Солонец солончаковатый |
14.24 |
58 : 6 : 36 |
6.4 |
162 |
|
I в. н. э. |
Каштановая несолонцеватая |
8.08 |
53 : 7 : 40 |
5.8 |
135 |
|
( 2000 лет назад) |
Каштановая солонцеватая |
8.97 |
57 : 5 : 38 |
7.8 |
149 |
|
|
||||||
|
Каштановая сильносолон& |
6.86 |
53 : 7 : 40 |
6.1 |
134 |
|
|
цеватая |
|
|
|
|
|
|
Солонец солончаковатый |
7.54 |
65 : 6 : 29 |
5.0 |
228 |
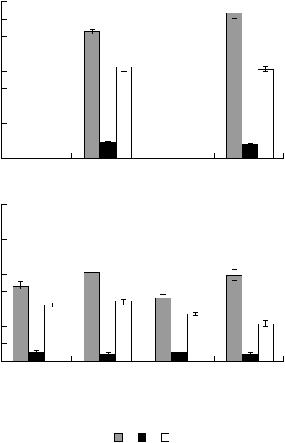
максимальные значения численности этих мик& |
Для сравнения исследованных почв в целом |
|
роорганизмов установлены в каштановой солон& |
были рассчитаны средневзвешенные значения |
|
цеватой почве, а минимальные – также в солон& |
(гор. А1 + В1 + В2) величин изученных микро& |
|
це. В гор. В2 степень солонцеватости почв на 80% |
биологических параметров. Показано, что сум& |
|
марная численность микроорганизмов в совре& |
||
обусловила изменения численности микроорга& |
||
менных почвах была больше (13–14 млн КОЕ/г |
||
низмов, довольствующихся низкими концентра& |
||
почвы), чем в подкурганных (7–9 млн КОЕ/г поч& |
||
циями элементов питания из рассеянного состоя& |
||
вы) (табл. 6). Уменьшение численности микроор& |
||
ния. Наибольшая численность этой группы за& |
||
ганизмов в подкурганных палеопочвах касалось |
||
фиксирована также в каштановой солонцеватой |
||
всех трофических групп. Так, численность мик& |
||
почве, а наименьшая – в каштановой сильносо& |
||
роорганизмов, использующих элементы питания |
||
лонцеватой. |
из рассеянного состояния, в современных почвах |
|
Различия почв по степени солонцеватости по& |
составила 7–8, в подкурганных – 4–5 млн КОЕ/г |
|
почвы; разлагающих гумус – 0.8–0.9 и 0.4–0.6; |
||
влияли и на биомассу темноокрашенного грибно& |
||
минерализующих растительные остатки – 5 и 2– |
||
го мицелия. Ее изменения в гор. А1 на 68%, а в |
||
3 млн КОЕ/г почвы соответственно (рис. 2). При |
||
гор. В1 на 81% зависели именно от этих различий |
сравнении современных почв выявлены досто& |
|
почв (табл. 5). |
верные различия: численность микроорганиз& |
ПОЧВОВЕДЕНИЕ № 7 2013
ХАРАКТЕРИСТИКА МИКРОБНЫХ СООБЩЕСТВ СОВРЕМЕННЫХ |
847 |
|
9 |
А |
|
|
|
|
|
|
|
|
8 |
|
|
|
|
7 |
|
|
|
|
6 |
|
|
|
|
5 |
|
|
|
|
4 |
|
|
|
|
3 |
|
|
|
почвы |
2 |
|
|
|
1 |
|
|
|
|
|
|
|
|
|
КОЕ/г |
0 |
|
|
Солонец |
Каштановая |
|
|||
|
|
|||
|
солонцеватая |
солончаковатый |
||
млн. |
9 |
Б |
|
|
8 |
|
|
|
|
|
|
|
|
|
|
7 |
|
|
|
|
6 |
|
|
|
|
5 |
|
|
|
|
4 |
|
|
|
|
3 |
|
|
|
|
2 |
|
|
|
|
1 |
|
|
|
|
0 |
|
|
|
|
Каштановая |
|
Каштановая |
|
|
несолонцеватая |
сильносолонцеватая |
||
|
Каштановая |
|
Солонец |
|
|
солонцеватая |
|
солончаковатый |
|
|
|
1 |
2 |
3 |
Рис. 2. Численность микроорганизмов различных трофических групп в современных (А) и погребенных (Б) почвах солонцовых комплексов (средневзвешенные значения величин в гор. А1 + В1 + В2). 1–3 – числен& ность микроорганизмов, выросших на: почвенном агаре (1), нитритном агаре (2), богатой органической среде (3).
мов, выросших на ПА, в профиле каштановой почвы на 20% меньше, а выросших на НА, на 10% больше, чем в солонце. Численность же микроор& ганизмов, выросших на БС, не имела достовер& ных различий в этих почвах (рис. 2А). В подкур& ганных палеопочвах наибольшая численность микроорганизмов, использующих элементы пи& тания из рассеянного состояния, установлена в каштановой солонцеватой почве и солонце, наи& меньшая (в 1.4 раза) – в каштановой сильносо& лонцеватой. Достоверно большая численность микроорганизмов, минерализующих гумус, обна& ружена в профиле каштановой несолонцеватой почвы (в 1.3 раза) по сравнению с другими палео& почвами, которые между собой достоверно не различались. Наибольшая численность микроор& ганизмов, разлагающих растительные остатки, выявлена в каштановых солонцеватой и несолон& цеватой почвах (различия между которыми не до&
стоверны), в 1.2 раза меньшая – в каштановой сильносолонцеватой и в 1.6 раза меньшая – в со& лонце (рис. 2Б). Следовательно, прослеживается обратная зависимость между численностью мик& роорганизмов, выросших на БС и степенью со& лонцеватости почв.
В ЭТС всех исследованных каштановых почв различия долей групп микроорганизмов, исполь& зующих как трудно&, так и легкодоступные орга& нические вещества, не превышали 2%; использу& ющих элементы питания из рассеянного состоя& ния – 4% (табл. 6). Четких различий между современной каштановой почвой и подкурган& ными палеопочвами не установлено как по ЭТС, так и по величинам БС/НА (5.7 и 5.8–7.8) и ин& дексу олиготрофности (139 и 134–149 соответ& ственно). Выявленная неоднородность в кашта& новых палеопочвах одного времени, вероятно, в значительной степени связана с различиями этих почв по степени солонцеватости. Существенные различия обнаружены для солонцов. При одина& ковой доли микроорганизмов, использующих гу& мусовые вещества (6%), доля микроорганизмов, разлагающих растительные остатки на 7% боль& ше, а довольствующих элементами питания из рассеянного состояния на 7% меньше в совре& менной почве по сравнению с палеосолонцом I в. н.э. Большие различия зафиксированы и по величинам БС/НА и индексу олиготрофности: в современном солонце они составили 6.4 и 162, в палеосолонце – 5.1 и 228 соответственно.
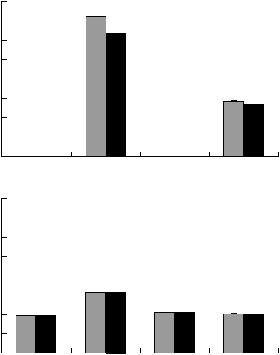
Наибольшая биомасса грибного мицелия об& наружена в профиле современной каштановой почвы: 73 мкг/г почвы, из них 83% приходились на темноокрашенные гифы (рис. 3А). В совре& менном солонце биомасса грибного мицелия бы& ла в 2.6 раза меньше, причем доля темноокрашен& ного мицелия возросла до 96%. В каштановых па& леопочвах I в. н. э. биомасса грибных гиф составила 20–32 мкг/г почвы, при этом 100% – темноокрашенные гифы. В этот же интервал укладывается и палеосолонец I в. н. э., его гриб& ная биомасса составила 20 мкг/г почвы при 100%&ном содержании темноокрашенных гиф. Наибольшая биомасса гиф обнаружена в каштано& вой солонцеватой почве (в 1.5–1.6 раза) по сравне& нию с другими палеопочвами, достоверные разли& чия между которыми не выявлены (рис. 3Б).
ЗАКЛЮЧЕНИЕ
Проведенные исследования микробных сооб& ществ современных и погребенных под курган& ной насыпью в I в. н. э. каштановых почв разной степени солонцеватости и солонцов показали, что закономерности изменения численности микроорганизмов различных трофических групп (довольствующихся элементами питания из рас& сеянного состояния, потребляющих гумус, ис&
ПОЧВОВЕДЕНИЕ № 7 2013
848
мкг/г почвы
ДЕМКИНА и др.
80 |
А |
|
|
|
|
70 |
|
|
60 |
|
|
50 |
|
|
40 |
|
|
30 |
|
|
20 |
|
|
10 |
|
|
0 |
Каштановая |
Солонец |
|
||
|
солонцеватая |
солончаковатый |
80 |
Б |
|
70 |
|
|
60 |
|
|
50 |
|
|
40 |
|
|
30 |
|
|
20 |
|
|
10 |
|
|
0 |
|
|
Каштановая Каштановая несолонцеватая сильносолонцеватая
Каштановая Солонец солонцеватая солончаковатый
микроорганизмов, минерализующих гумус, в гор. В1 на численности микроорганизмов, ис& пользующих легкодоступные растительные остатки, в гор. В2 на численности микроорганиз& мов, довольствующих низкими концентрациями элементов питания из рассеянного состояния. В гор. А1 и В1 наименьшая численность этих мик& роорганизмов зафиксирована также в солонце. При сравнении изученных почв в целом по сред& невзвешенным в профиле величинам микробио& логических параметров прослеживается обратная зависимость между численностью микроорганиз& мов, использующих легкодоступное органическое вещество (растительные остатки) и степенью со& лонцеватости подкурганных палеопочв I в. н. э. В численности других трофических групп микроор& ганизмов закономерности, связанные со степе& нью солонцеватости почв, не выявлены. Наи& большая биомасса грибного мицелия обнаружена в каштановой солонцеватой палеопочве (в 1.5– 1.6 раза) по сравнению с другими палеопочвами, которые между собой достоверно не различались.
Таким образом, пространственное варьирова& ние микробиологических параметров, связанное с комплексностью почвенного покрова сухостеп& ной зоны, находится в пределах кратности разли& чий 1.2–2.8, что несомненно следует учитывать при исследовании их временноdй динамики.

 2
2  1
1
Рис. 3. Биомасса и структура грибного мицелия в со& временных (А) и погребенных (Б) почвах солонцовых комплексов (средневзвешенные значения величин в гор. А1 + В1 + В2). Обозначения: 1 – суммарная био& масса грибного мицелия; 2 – биомасса темноокра& шенного мицелия.
пользующих легкодоступное органическое веще& ство) и биомассы грибного мицелия по профилю этих почв во многом сходны и не имеют суще& ственных отличий. На основе дисперсионного анализа дана количественная оценка влияния со& лонцового процесса на пространственные изме& нения микробиологических параметров исследо& ванных почв. Сравнение каштановой почвы и со& лонца по горизонтам выявило достоверные различия как в современных почвах, так и в под& курганных палеопочвах I в. н. э. Как правило, в каштановых почвах величины микробиологиче& ских параметров были в 1.2–2.8 раза больше, чем в солонцах. Проведенная оценка влияния степе& ни солонцеватости на изменения величин микро& биологических параметров в различных горизон& тах подкурганных почв показала, что это влияние проявилось во всем исследованном почвенном профиле, но в каждом горизонте достоверно от& разилось только на численности какой&то одной трофической группы: в гор. А1 на численности
СПИСОК ЛИТЕРАТУРЫ
1.Ананьева Н.Д., Васильева Г.К. Роль микробиологи& ческого фактора в разложении 3,4&дихлоранилина в почвах // Почвоведение. 1985. № 5. С. 57–64.
2.Аринушкина Е.В. Руководство по химическому анализу почв. М.: Изд&во Моск. ун&та, 1970. 487 с.
3.Всеволодова Перель Т.С., Колесников А.В. Количе& ственная характеристика мезофауны почв солон& цового комплекса полупустыни Северного Прика& спия // Почвоведение. 2010. № 11. С. 1381–1386.
4.Демкин В.А. Палеопочвоведение и археология: ин& теграция в изучении истории природы и общества. Пущино: ОНТИ ПНЦ РАН, 1997. 213 с.
5.Демкин В.А., Борисов А.В., Демкина Т.С. и др. Вол& го&Донские степи в древности и средневековье (по материалам почвенно&археологических исследо& ваний). Пущино: SYNCHROBOOK, 2010. 120 с.
6.Демкин В.А., Демкина Т.С., Алексеев А.О. и др. Па& леопочвы и климат степей Нижнего Поволжья в I– IV вв. н.э. Пущино: ОНТИ ПНЦ РАН, 2009. 96 с.
7.Демкина Т.С., Борисов А.В., Демкин В.А. Микроб& ные сообщества палеопочв археологических па& мятников пустынно&степной зоны // Почвоведе& ние. 2000. № 9. С. 1117–1126.
8.Демкина Т.С., Борисов А.В., Демкин В.А. Микро& биологические исследования подкурганных па& леопочв пустынно&степной зоны Волго&Донского междуречья // Почвоведение. 2004. № 7. С. 853– 859.
ПОЧВОВЕДЕНИЕ № 7 2013
ХАРАКТЕРИСТИКА МИКРОБНЫХ СООБЩЕСТВ СОВРЕМЕННЫХ |
849 |
9. Демкина Т.С., Борисов А.В., Демкин В.А. Палеопоч& вы и природная среда Северных Ергеней в эпохи энеолита и бронзы (IV–II тыс. до н.э.) // Почвове& дение. 2003. № 6. С. 655–669.
10. Демкина Т.С., Мирчинк Т.Г. Определение грибной биомассы в почвах методом мембранных фильтров // Микология и фитопатология. 1983. Т. 17. Вып. 6. С. 517–520.
11. Демкина Т.С., Стретович И.В., Демкин В.А. Про& странственная изменчивость микробных сооб& ществ современных и погребенных почв в бассей& не р. Сакарка (Приволжская возвышенность) // Почвоведение. 2010. № 5. С. 621–631.
12.Демкина Т.С., Хомутова Т.Э., Борисов А.В., Демкин В.А. Микробиологические исследования подкурганных палеопочв в долине реки Иловли // Мат&лы по археологии Волго&Донских степей. Волгоград: Изд&во ВолГУ, 2004. Вып. 2. С. 87–95.
13.Демкина Т.С., Хомутова Т.Э., Каширская Н.Н. и др.
Микробиологические исследования палеопочв ар& хеологических памятников степной зоны // Поч& воведение. 2010. № 2. С. 213–220.
14.Жданова Н.Н., Костюк М.Д., Северинова А.Е. Дыха& ние некоторых темноокрашенных гифомицетов в условиях продолжительного голодания // Изве& стия АН СССР. Сер. биологическая. 1982. № 6. С. 912–922.
15.Жданова Н.Н., Походенко В.Д. О возможном уча& стии меланинового пигмента в защите грибной
клетки от высыхания // Микробиология. 1973.
Т. 42. Вып. 5. С. 848–853.
16.Звягинцев Д.Г., Асеева И.В., Бабьева И.П., Мирчинк Т.Г. Методы почвенной микробиологии и биохимии. М.: Изд&во Моск. ун&та, 1980. 224 с.
17.Казеев К.Ш., Кременица А.М., Колесников С.И. и др.
Биологические свойства почв каштаново&солон& цовых комплексов // Почвоведение. 2005. № 4. С. 464–474.
18. Никитин Д.И., Никитина Э.С. Процессы самоочи& щения окружающей среды и паразиты бактерий (род Bdellovibrio). М., 1978. 205 с.
19. Теппер Е.З. Микроорганизмы рода Nocardia и раз& ложение гумуса. М.: Наука, 1976. 199 с.
20.Хитров Н.Б. Связь почв солонцового комплекса Северного Прикаспия с микрорельефом // Почво& ведение. 2005. № 3. С. 271–284.
21.Bloomfield B.J., Alexander M. Melanins and resistance of fungi to lysis // J. of Bacteriology. 1967. V. 93. № 4.
P.1276–1280.
22.COHORT Software. 1986, 1990. Costat version 4.21, Berkely. CA, USA.
23.Demkina T.S., Khomutova T.E., Kashirskaya N.N., Demkina E.V., Stretovich I.V., El Registan G.I., Demkin V.A. Age and activation of microbial commu& nities in soils under burial mounds and in recent surface soils of steppe zone // Eurasian Soil Science. 2008.
V.41. № 13. P. 1439–1447.
6 ПОЧВОВЕДЕНИЕ № 7 2013

ПОЧВОВЕДЕНИЕ, 2013, № 7, с. 850–859
БИОЛОГИЯ
ПОЧВ
УДК 631.461:631.416.1:631.417.1
ВЛИЯНИЕ ВЫСУШИВАНИЯ ОБРАЗЦОВ ГОРНО ЛУГОВЫХ АЛЬПИЙСКИХ ПОЧВ НА ПРОЦЕССЫ ТРАНСФОРМАЦИИ СОЕДИНЕНИЙ АЗОТА И УГЛЕРОДА*
© 2013 г. М. И. Макаров1, О. С. Мулюкова1, Т. И. Малышева1, О. В. Меняйло2
1Факультет почвоведения МГУ им. М.В. Ломоносова, 119991, Москва, Ленинские горы 2Институт леса им. В.Н. Сукачева Сибирского отделения РАН, 660036, Красноярск, Академгородок e*mail: mikhail_makarov@mail.ru
Поступила в редакцию 07.08.2012 г.
Высушивание образцов горно$луговых альпийских почв, для которых в естественных условиях ха$ рактерна постоянно высокая влажность, принципиально изменяет концентрацию лабильных со$ единений азота и углерода и закономерности их микробной трансформации. При высушивании об$ разцов почв в них в 4–5 раз увеличивается содержание экстрагируемых органических соединений азота и углерода и неорганических соединений азота, а количество азота и углерода микробной био$ массы уменьшается в 2–3 раза. После увлажнения высушенной почвы в ней начинается процесс восстановления запасов азота и углерода микробной биомассы, однако после двух недель инкуба$ ции эти показатели все еще в 1.5–2 раза уступают исходным значениям, характерным для естествен$ ной почвы. Восстановление микробного сообщества в образцах предварительно высушенных почв происходит в отсутствии недостатка лабильных органических соединений и сопровождается их ак$ тивной минерализацией и слабой ассимиляцией аммонийного азота микроорганизмами.
Ключевые слова: горно$луговые почвы, углерод, азот, микробная биомасса, нитрификация, минера$ лизация, иммобилизация.
DOI: 10.7868/S0032180X1307006X
ВВЕДЕНИЕ
Влияние высушивания и последующего увлажнения почвы на ее свойства и активность процессов микробной трансформации соедине$ ний азота и углерода изучается на протяжении не$ скольких десятилетий [11, 14, 24, 26, 32]. Это свя$ зано с тем, что, с одной стороны, такое сочетание резко меняющихся условий увлажнения почвы широко распространено в природе, а с другой, со$ вокупность процессов высушивания и увлажне$ ния почвы сопровождает стандартную процедуру анализа воздушно$сухих образцов.
Известно, что высушивание и последующее увлажнение почвы приводит к изменению ее ла$ бильных свойств. Высушивание почвы и хране$ ние в воздушно$сухом состоянии до анализа уменьшает ее ферментативную активность [3, 26]. При высыхании почвы происходит гибель микро$ организмов, и уменьшается концентрация N и C микробной биомассы (N микр и C микр) [20], а ко$ личество N и C экстрагируемых органических со$ единений (N орг и C орг) и N неорганических со$
*Работа выполнена при финансовой поддержке РФФИ (проект 10$04$00780).
единений (N–NH+4 и N–NO 3− ) возрастает [32]. После увлажнения сухой почвы устойчивые к вы$ сушиванию микроорганизмы получают доступ к легкодоступному субстрату, в результате чего рез$ ко возрастает ее биологическая активность, выра$ жающаяся в увеличении скорости минерализа$ ции C и N [11, 14, 24, 32].
В связи с этим в методических рекомендациях по определению лабильных свойств почв (микро$ биологических, биохимических, форм соедине$ ний азота и углерода) указывается на необходи$ мость анализа свежих образцов и максимально возможного сокращения времени их хранения при естественной влажности и температуре +4 или –20°C [9, 10, 23, 30]. Однако в практике проведения научных исследований эти условия не всегда выполняются, и исследователи исходят из технической простоты осуществления процес$ са хранения почвенных образцов, включая их хранение в воздушно$сухом состоянии. В частно$ сти, в одной из недавних публикаций при прове$ дении массовых исследований ферментативной активности почв предпочтение отдается хране$ нию образцов в воздушно$сухом состоянии при
850