Материал: В-Л-Быков-Цитогогия_и_общая_гистология
цепторами, не получая необходимой для роста стимуляции, подвергаются апоптозу. Соответственно, антитела, которые будут продуцироваться плазматическими клетками - потомками В-лимфоцитов - будут постепенно приобретать все более высокую аффинность (процесс созревания аффинности) и повышенную способность к нейтрализации или элиминации антигена. Эффективной селекции способствует процесс соматической гипермутации, который вносит еще большее разнообразие в обширный репертуар рецепторов, обусловленный реаранжировкой генома В-лимфоцитов в красном костном мозге. Этот процесс запускается в результате взаимодействия активированных В-лимфоцитов с Т-лимфоцитами.
Плазмобласты и плазмоциты. Плазмобласты отличаются от иммунобластов усиленным развитием грЭПС и комплекса Гольджи, синтезом и секрецией иммуноглобулинов. По мере их преобразования в плазматические клетки синтетические процессы еще более усиливаются. Происходит дальнейшее увеличение доли объема цитоплазмы, занятой грЭПС. На светооптическом уровне выявляется усиление базофилии по всей цитоплазме, за исключением светлого околоядерного "дворика", соответствующего месту расположения комплекса Гольджи и центриолей. Ядро уменьшается в размерах, занимает эксцентричное положение в клетке, хроматин конденсируется с образованием характерной картины "спиц колеса" (см. рис. 8-8).
В процессе развития плазматических клеток происходит потеря части специфических маркеров, свойственных В-лимфоцитам (например, связанных с мембраной иммуноглобулинов, репепторов СЗ-компонента комплемента, Fcфрагмента иммуноглобулинов, CD19 и CD21).
Топография дифференцирующихся плазматических клеток.
Образование плазматических клеток может происходить в периферических лимфоидных органах или (при мшрации активированных клеток с током крови в периферические ткани) - в собственной пластинке слизистых оболочек, строме желез. Оно осуществляется также в красном костном мозге.
Часть В-иммунобластов превращается в долгоживущие В-клетки памяти с высокоаффинными поверхностными рецепторами, функция которых заключается в обеспечении быстрой реакции на повторный контакт с антигенами.
РАЗВИТИЕ Т-ЛИМФОЦИТОВ
Последовательность стадий антиген-независимого развития Т-
лимфоцитов представлена на рис. 9-7(1).
- 271 -
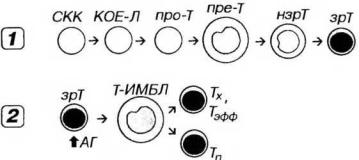
Рис. 9-7. Развитие Т-лимфоцитов. 1 - антиген-независимое, 2 - антигензависимое Про-Т - про-Т-л и мфоцит (протимоцит), пре-Т - пре-Т-лимфоцит (претимоцит), нзрТ - незрелый Т-лимфоцит, зрТ - зрелый Т-лимфоцит, зрТ - зрелый Т-лимфоцит, Т-ИМБЛ - Т-иммунобласт, Тэфф - Т-лимфоцит-эффектор, Тх - Т-хелпер, Тп - Т-клетка памяти, остальные обозначения - как на рис. 9-6.
Протимоцит (про-Т-лимфоцит) образуется в красном костном мозге из КОЕ-Л и соответствует стадии, предшествующей реаранжировке генома.
Претимоцит - наиболее ранняя стадия развития Т-лимфоцитов в тимусе после миграции из красного костного мозга. В нем начинается реаранжировка генома, однако экспрессия Т-клеточных рецепторов (ТКР) на поверхности клетки отсутствует. На плазмолемме имеются поверхностные маркеры, свойственные незрелым клеткам.
Незрелые Т-лимфоциты и зрелые Т-лимфоциты - последовательные стадии, идущие за претимоцитами. Эти клетки претерпевают реаранжировку генома с формированием разнообразных специфических антиген-распознающих ТКР, которые экспрессируются на их поверхности. На плазмолемме появляется ряд маркеров, типичных для зрелых Т-лимфоцитов и необходимых для их взаимодействия с другими клетками. Одновременно утрачиваются маркеры, свойственные незрелым клеткам. При этом их фенотип изменяется следующим образом:
ТКР-/СD3-/СD4-/СD8- -> ТКР+/СD3+/СD4+/С08+ ->
ТКР+/СD3+/СD4+/СD8или ТКР+/СD3+/СD4-/С08+ .
Морфологически претимоциты соответствуют лимфобластам, незрелые Т-лимфоциты - средним лимфоцитам, а зрелые Т-лимфо-циты - малым лимфоцитам.
- 272 -
Развитие Т-лимфопитов в тимусе регулируется их контактными взаимодействиями с эпителиальными клетками, образующими строму этого органа, а также разнообразными гемопоэтинами, продуцируемыми, в частности, этими клетками. К ним относятся различные КСФ, ИЛ-1, ИЛ-6, а также ряд специфических тимусных факторов - тимозин, тимопоэтин, тимусный сывороточный фактор и др.
Последовательность стадий антиген-зависимого развития Т-
лимфоцитов представлена на рис. 9-7(2).
Покидая тимус, наивные (зрелые) Т-лимфоциты с током крови мигрируют в Т-зависимые зоны периферических органов кроветворения и иммуногенеза. В этих органах они встречаются с антигенами, которые им представляют АПК после процессинга, и взаимодействуют с Т-хелперами.
Взаимодействуя с антигеном, который находится в комплексе с молекулами МНС, а также получая дополнительные сигналы при адгезионных контактах и воздействии цитокинов, Т-лимфоциты активируются, подвергаются бласт-трансформации - превращаются в Т-иммунобласты. Последние пролиферируют и дифференцируются, формируя крупные клоны эффекторных и регуляторных клеток (см. главу 8). Часть Т-лимфоцитов превращается в долгоживущие Т-клетки памяти с фенотипом СD45RO+ и усиленной экспрессией ТКР и ряда маркеров, которые придают им высокую чувствительность к повторному воздействию данного антигена.
РАЗВИТИЕ NK-КЛЕТОК
NK-клетки происходят из костномозгового предшественника, причем их развитие не связано с образованием Т- и В-лимфоцитов. Полагают, что наряду с костным мозгом, они могут развиваться также и в тимусе. После выхода в кровь NK-клетки циркулируют в ней или мигрируют в селезенку; в лимфатических узлах содержатся лишь единичные NK-клетки. Их дозревание происходит в тканях под влиянием малоизученных факторов микроокружения. Механизмы, регулирующие рециркуляцию NK-клеток и их миграцию в селезенку, остаются малоизученными; но всей видимости, они опосредуются адгезивными взаимодействиями между NK-клетками и эндотелием сосудов.
- 273 -
СТРОЕНИЕ И ГИСТОФИЗИОЛОГИЯ МИЕЛОИДНОЙ И ЛИФОИДНОЙ ТКАНЕЙ
Миелоидная и лимфоидная ткани являются кроветворными тканями, которые представляют собой особые виды соединительных тканей, или тканей внутренней среды (см. главу 6). В состав каждой из этих тканей входят два компонента:
(1)форменные элементы крови на различных стадиях развития (описание см. выше);
(2)ретикулярная ткань.
СТРОЕНИЕ И ГИСТОФИЗИОЛОГИЯ РЕТИКУЛЯРНОЙ ТКАНИ
Ретикулярная ткань относится к соединительным тканям со специальными свойствами и обеспечивает развитие форменных элементов крови. Она является главным элементом, образующим структурную основу (строму) кроветворных тканей (миелоидной и лимфоидной) во всех органах кроветворения и иммуногенеза. Лишь лимфоидная ткань тимуса служит исключением из общего правила, поскольку в ней место ретикулярной ткани занимает специализированная эпителиальная ткань.
Функции ретикулярной ткани. Наиболее общая функция ретикулярной ткани - обеспечение процессов кроветворения путем создания необходимого микроокружения для развивающихся клеток крови. Она включает ряд более частных функций - опорную, трофическую, секреторную, фагоцитарную и (в периферических органах кроветворения и иммуногенеза) антигенпредставляющую.
Компонентами ретикулярной ткани являются клетки и межклеточное вещество (рис. 9-8).
Клетки ретикулярной ткани подразделяются на фиксированные - ретикулярные клетки (ведущий компонент) и свободные - макрофаги и дендритные антиген-представляющие клетки.
Ретикулярные клетки - крупные отростчатые фибробластоподобные клетки, формирующие сеть, которая пронизывает кроветворные ткани и образует их структурную основу. Они характеризуются большим округлым центрально расположенным светлым (с преобладанием эухроматина) ядром с крупным ядрышком, слабооксифильной цитоплазмой, в которой при электронно-микроскопическом исследовании обнаруживаются умеренно развитые органеллы, хорошо выраженный
- 274 -
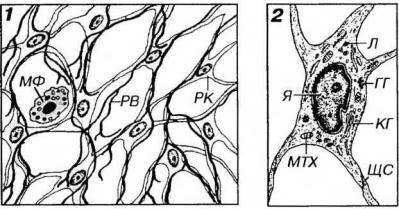
цитоскелет, включения гликогена. Ретикулярные клетки связаны друг с другом посредством щелевых соединений; к их поверхности прилежат ретикулярные волокна, которые частично вдавливаются в их цитоплазму.
Адвентициальные клетки - одна из разновидностей ретикулярных клеток в миелоидной ткани, которые снаружи вплотную прилежат к эндотелию венозных синусов красного костного мозга, образуя их наружную оболочку - адвентицию. Эти клетки, по-видимому, обладающие достаточно высоким уровнем дифференцировки, не следует смешивать с малодифференцированными клетками рыхлой волокнистой соединительной ткани, носящими то же название (см. главу 10). Адветициальные клетки регулируют миграцию зрелых форменных элементов из миелоидной ткани в кровь, создавая своеобразный барьер на их пути. Близкую функцию контроля миграции форменных элементов крови, возможно, выполняют и адвентициальные (ретикулярные) клетки, охватывающие венозные синусы в селезенке.
Ретикулярные клетки и адипоциты. Высказывается предположение, что малодифференцированные предшественники ретикулярных клеток, накапливая липиды, могут (подобно малодифференцированным фибробластам - см. главы 10 и 11) превращаться в жировые клетки (адипоциты), особенно многочисленные в миелоидной ткани. Согласно другим взглядам, адипоциты представляют собой самостоятельный элемент, входящий в состав стромы кроветворных органов.
Рис. 9-8. Ретикулярная ткань. 1 - общий вид ткани на гистологическом препарате; 2 - ультраструктурная организация ретикулярной метки. РК - ретикулярные клетки, РВ - ретикулярные волокна, МФ - макрофаг, ЩС - щелевое соединение, Я - ядро, КГ - комплекс Гольджи, МТХ - митохондрия, Л - лизосомы, ГГ - гранулы гликогена.
- 275 -