Материал: В-Л-Быков-Цитогогия_и_общая_гистология
ТРОМБОЦИТОПОЭЗ
Тромбоцнтопоэз - процесс образования и созревания тромбоцитов, происходящий в миелоидной ткани. Тромбоциты образуются в результате процесса частичной фрагментации цитоплазмы гигантских клеток костного мозга - мегакариоцитов. Ход развития мегакариоцитов из стволовой клетки крови описывается последовательностью:
СКК -> КОЕ-ГЭММ -> КОЕ-Мег -> мегакариобласт -> мегакариоцит.
СТРУКТУРНО-ФУНКЦИОНАЛЬНЫЕ ИЗМЕНЕНИЯ МЕГАКАРИОЦИТА В ХОДЕ ЕГО СОЗРЕВАНИЯ
Мегакариоцит при созревании из мегакариобласта становится крупнее, достигая 20-50 мкм (по некоторым сведениям - до 100 мкм) в диаметре; его ядро и цитоплазма претерпевают выраженные изменения.
Дифференцировка ядра включает активную репликацию ДНК без митоза. Полиплоидные клетки в дальнейшем претерпевают эндомитоз с образованием многочисленных связанных перемычками долей ядра, в котором общее содержание ДНК соответствует 4-128n (наиболее часто - 16 или 32n). Хроматин постепенно конденсируется, ядрышко, как правило, не выявляется. Размеры клетки обычно соответствуют степени ее полиплоидизации.
Дифференцировка цитоплазмы мегакариоцитов начинается только по завершении репликации ДНК. Наиболее заметными ее проявлениями служат:
(1)разделение цитоплазмы на три зоны: околоядерную,
промежуточную и краевую (периферическую).
Околоядерная зона содержит элементы грЭПС, хорошо развитый комплекс Гольджи, митохондрии и центриоли.
Промежуточная зона - наибольшая по ширине, содержит гранулы и систему мембранных демаркационных каналов (см. ниже).
Краевая (периферическая) зона свободна от большинства органелл и гранул, в ней в значительном количестве сосредоточены элементы цитоскелета (преимущественно актиновые микрофиламенты и ассоциированные с ними белки). Она пересекается демаркационными каналами, связанными с поверхностью клетки.
(2)образование и накопление гранул, характерных для тромбоцитов и содержащих типичные для них белки (см. главу 7); гранулы отсутствуют в узкой периферической зоне цитоплазмы;
-261 -
(3)формирование системы мембран (демаркационных каналов),
разрезающих цитоплазму мегакариоцитов на территории размером 2-4 мкм, соответствующие границам будущих тромбоцитов и содержащие гранулы. Демаркационные каналы возникают предположительно в результате инвагинации плазмолеммы и сообщаются с межклеточным пространством (согласно другим взглядам, они происходят из аЭПС или путем слияния пузырьков, продуцируемых комплексом Гольджи);
(4)образование филоподий (протромбоцитов) - узких длинных (2.5x120
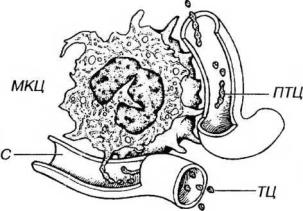
мкм) лентовидных отростков мегакариоцитов, которые через поры эндотелия синусов красного костного мозга проникают в их просвет, где распадаются на отдельные тромбоциты (рис. 9-4). Каждый зрелый мегакариоцит образует несколько тысяч (до 8000) тромбоцитов. В некоторых случаях мегакариоциты способны отделять в просвет сосудов костного мозга крупные фрагменты цитоплазмы.
Рис. 9-4. Выделение тромбоцитов мегакариоцитом в кровоток. Мегакариоцит (МКЦ) образует узкие лентовидные отростки - филоподии, или протромбоциты (ПТЦ). которые через поры эндотелия синусов (С) красного костного мозга проникают в их просвет, где распадаются на отдельные тромбоциты (ТЦ).
Неэффективный тромбоцитопоэз - процесс, при котором часть цитоплазмы мегакариоцитов (потенциально способная образовывать тромбоциты) остается в миелоидной ткани, не формируя филоподий. В норме активность этого процесса невелика, однако в патологических условиях он может приобретать значительные масштабы вследствие аномалий конечных стадий развитая мегакариоцитов или их удаления от стенки синусов.
- 262 -
Остаточные мегакариоциты - клетки после полного выделения тромбоцитов, в которых сохраняется лишь узкий ободок цитоплазмы вокруг ядра (околоядерная зона). Они составляют около 10% мегакариоцитов в миелоидной ткани, подвергаются дегенерации, фагоцитируются и замещаются новыми. Высказывается, однако, предположение о способности остаточных мегакариоцитов к восстановлению цитоплазмы и продукции нового поколения тромбоцитов.
Тромбоциты внекостномозгоеого происхождения продуцируются мегакариоцитами, которые через стенку синусов красного костного мозга целиком мигрировали в их просвет и после циркуляции в крови "застряли" в узких сосудах (например, почки, селезенки, печени и, особенно часто - легкого). Предполагают, что каждую минуту в легкое таким путем попадают десятки тысяч мегакарицитов. Благодаря их деятельности содержание тромбоцитов в крови, полученной из легочных вен, значительно выше, чем во взятой из легочной артерии. Более того, высказывается мнение, что продукция тромбоцитов мегакариоцитами в легких достаточна для поддержания их нормального уровня в крови и, возможно, даже превышает таковую в миелоидной ткани.
Цикл развития от стволовой клетки до формирования тромбоцитов
занимает около 10 сут. Тромбоцитопоэз контролируется рядом гуморальных факторов, из которых наибольшее значение имеют КСФ-Мег (стимулирует пролиферацию КОЕ-Мег) и тромбопоэтин (ускоряет созревание мегакариоцитов). Эти гуморальные факторы поддерживают скорость продукции тромбоцитов на необходимом для орга-низма уровне, быстро повышая ее в случае возникающей потребности. Так, через несколько дней после кровопотери с развитием тромбоцитопении содержание мегакариоцитов в миелоидной ткани увеличивается в 3-4 раза, а уровни тромбоцитов в крови - в 1.5-2 раза по сравнению с нормой.
ГРАНУЛОЦИТОПОЭЗ
Гранулоцитопоэз - образование и дифференцировка гранулоцитов - происходит в красном костном мозге. Исходным источником развития всех гранулоцитов служит СКК, которая дает начало КОЕ-ГЭММ. В отношении
ближайших потомков последней, однако, в литературе имеются некоторые разногласия, касающиеся уровня, на котором развитие гранулоцитов разделяется на самостоятельные клеточные линии (нейтрофилов, базофилов и эозинофилов). В соответствии с большинством современных источников, это происходит непосредственно после
- 263 -
уровня КОЕ-ГЭММ с формированием раздельных КОЕ для каждой линии - КОЕ-ГМ (дающей начало КОЕ-Г [нейтрофильных] и КОЕ-Мо), КОЕ-Баз и КОЕЭо (см. рис. 9-2). На некоторых схемах кроветворения это разделение обозначено после стадий КОЕ-ГМ, миелобласта и даже промиелоцита.
Последовательность начальных этапов развития гранулоцитов:
(а) нейтрофильных: СКК -> КОЕ-ГЭММ -> (б) базофильных: СКК -> КОЕ-ГЭММ (в) эозинофильных: СКК -> КОЕ-ГЭММ ->
КОЕ-ГМ -> КОЕ-Г(Н); -> КОЕ-Баз; КОЕ-Эо.
Последующие стадии развития гранулоцитов протекают для всех трех типов клеток однотипно (рис. 9-5):
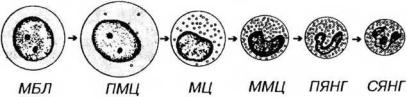
миелобласт -> промиелоцит -> миелоцит -> метамиелоцит -> палочкоядерный гранулоцит -> сегментоядерный гранулоцит.
Рис. 9-5. Гранулоцитопоэз (на примере развития нейтрофильных гранулоцитов). МБЛ - миелобласт, ПМЦ - промиелоцит, МЦ - миелоцит, ММЦ - метамиелоцит, ПЯНГ - палочкоядерный нейтрофильный гранулоцит, СЯНГ - сегментоядерный нейтрофильный гранулоцит.
При анализе развития базофильных и эозинофильных гранулоцитов стадии палочкоядерной и сегментоядерной клеток обычно не разделяют.
Процесс дифференцировки предшественников гранулоцитов в зрелые клетки включает (см. рис. 9-5):
(1)уменьшение размеров клетки;
(2)снижение, а в дальнейшем (со стадии метамиелоцита) - утрату способности к делению;
(3)изменение формы ядра - от округлой до бобовидной и палочковидной, его сегментация; нарастание конденсации ядерного хроматина;
- 264 -
(4) выработку и накопление гранул в цитоплазме;
(5)изменение состава гранул с постепенным увеличением доли специфических гранул и снижением содержания азурофильных;
(6)нарастание подвижности клетки, обусловленное перестройкой цитоскелета с увеличением содержания актиновых микрофиламентов;
(7)приобретение разнообразных рецепторов плазмолеммы, опосредующих адгезивные взаимодействия с другими клетками и компонентами межклеточного вещества и обеспечивающих важнейшие функции клеток - фагоцитоз, хемотаксис, секреторные реакции.
Промиелоцит - крупная (диаметр - 16-24 мкм) клетка с развитой слабобазофильной цитоплазмой и большим круглым светлым ядром, содержащим мелкодисперсный хроматин и 1-2 ядрышка. Многочисленные полисомы, цистерны грЭПС и крупный комплекс Голыши обеспечивают образование первичных (азурофильных) гранул (лизосом) диаметром 500 нм, содержание которых по мере созревания промиелоцита увеличивается. К концу этой стадии образование азурофильных гранул завершается. Поэтому, поскольку промиелоцит активно делится, на последующих стадиях содержание этих гранул непрерывно снижается. К концу развития в промиелопите появляются единичные вторичные (специфические - нейтрофильные, базофильные или эозинофильные) гранулы.
Миелоцит обычно меньших размеров, чем промиелопит (диаметр - 1016 мкм). Характеризуется односторонним уплощением или небольшой инвагинацией эксцентрично расположенного ядра с более крупными гранулами гетерохроматина; ядрышки исчезают. Цитоплазма содержит первичные гранулы (количество которых постепенно падает) и вторичные (специфические) гранулы (число которых непрерывно возрастает благодаря деятельности синтезирующих их грЭПС и комплекса Гольджи). К концу стадии миелоцита вторичных гранул становится больше, чем первичных, происходит накопление гликогена. Миелоцит - последняя митотически активная клетка линии гранулоцитов.
Метамиелоцит - меньших размеров (по сравнению с миелоцитом) и отличается от него более заметной инвагинацией ядра, которое принимает бобовидную форму и уплотняется. По ходу созревания метамиелоцита, а позднее на стадиях палочко- и сегментоядерного гранулоцита происходят дальнейшие изменения ядра и цитоплазмы.
Ядро измеггяет форму - из бобовидного становится подковообразным, а затем палочковидным с последующим формированием перетяжек, разделяющих его на сегменты; конденсация хроматина прогрессивно нарастает.
- 265 -