Материал: В-Л-Быков-Цитогогия_и_общая_гистология
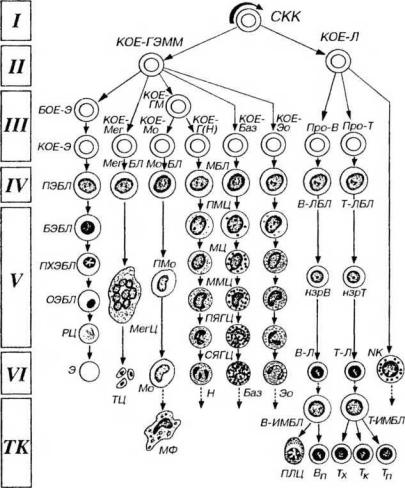
Рис 9-2. Схема кроветворения. I-VI - классы кроветворных клеток, СКК - стволовая клетка крови, КОЕ - колониеобразующая единица (родоначальная клетка): КОЕ-ГЭММ - КОЕ гранулоцитов, эритроцитов, моноцитов и мегакариоцитов, КОЕ-Э - КОЕ эритроцитов, КОЕ-Мег - КОЕ мегакариоцитов, КОЕ-ГМ - КОЕ гранулоцитов (нейтрофильных) и моноцитов, КОЕ-Г(Н) - КОЕ гранулоцитов (нейтрофильных), КОЕ-Мо - КОЕ моноцитов, КОЕ-Баз - КОЕ базофилов, КОЕ-Эо - КОЕ эозинофилов, КОЕ-Л - КОЕ лимфоцитопоэза, БОЕ-Э - бурст-образующая единица, про-В - про-В- лимфоцит, про-Т - про-Т- лимфоцит (протимоцит), ПЭБЛ - проэритробласт, БЭБЛ - базофильный эритробласт, ПХЭБЛ - полихроматофильный эритробласт, ОЭБЛ - оксифильный (ортохроматофильный) эритробласт, РЦ - ретикулоцит, Э - эритроцит. МегБЛ - мегакариобласт, МегЦ - мегакариоцит, ТЦ - тромбоциты, МоБЛ - моноцитобласт, ПМо - промоноцит, Мо - моноцит, МФ - макрофаг, МБЛ - миелобласты, ГШЦ - промиелоциты, МЦ - миелоциты, ММЦ - метамиелоциты, ПЯГЦ – палочкоядерные
- 256 -
гранулоциты, СЯГЦ - сегментоядерные гранулоциты (нейтрофильный - Н, базофильный - Баз, эозинофильный - Эо). В-ЛБЛ - В-лимфобласт, нзрВ - незрелый В-лимфоцит, В-Л - В-лимфоцит (зрелый), В-ИМБЛ - В-иммунобласт, ПЛЦ - плазмоцит, Вп - В-клетка памяти, Т-ЛБЛ - Т-лимфобласт , нзрТ - незрелый Т-лимфоцит, Т-Л - Т-лимфоцит (зрелый), Т-ИМБЛ - Т-иммунобласт, Тх - Т-хелпер, Тк - Т-киллер, Тп - Т-клетка памяти, ГТК - ЫК-клетка. Миграция зрелых клеток из крови в периферические ткани (ТК) обозначена пунктирными стрелками; пути рециркуляции лимфоцитов не отмечены.
Форменные элементы крови в тканях. Из зрелых форменных элементов лишь эритроциты и (частично) тромбоцита выполняют свои функции исключительно в кровотоке, лейкоциты же реализуют их после миграции в ткани. Часть клеток при этом подвергается дальнейшим преобразованиям (например, моноциты превращаются в макрофаги и дендритные АПК, лимфоциты под действием антигенной стимуляции подвергаются бласттрансформации и дальнейшей дифференцировке, основная часть В-лимфоцитов дифференцируется в плазматические клетки).
ЭРИТРОПОЭЗ
Эритропоэз (эритроцитопоэз) - процесс образования и созревания эритроцитов, происходящий в миелоидной ткани. Ход развития эритроцитов из стволовой клетки крови описывается последовательностью:
СКК -> КОЕ-ГЭММ -> БОЕ-Э -> КОЕ-Э -> проэритробласт > базофильный эритробласт -> полихроматофнльный эритробласт -> оксифильный (ортохроматофильный) эритробласт -> ретикулоцит -> эритроцит.
Эритрон - эритроидный дифферон, представляющий собой совокупность указанных форм - от эритроидных родоначальных клеток до зрелых эритроцитов (включая циркулирующие в крови).
БОЕ-Э и КОЕ-Э. БОЕ-Э - бурст-образующая единица (от англ. burst - взрыв) - названа так по своей способности быстро (взрывоподобно) образовывать на полутвердой среде колонию эритроидных клеток численностью в несколько сотен элементов. Она отличается от развивающейся из нее КОЕ-Э более высокой пролиферативной активностью, высокой чувствительностью к ИЛ-3 и низкой - к эритропоэтину.
Терминология, используемая для наименования дальнейших стадий развития клеток эритроидного ряда (следующих за КОЕ-Э) неоднозначна. Термины эритробласт, нормобласт и нормоцит применяются
- 257 -
разными авторами для обозначения одних и тех же клеточных форм. В настоящей книге использованы наименования указанных клеток, получившие наиболее широкое распространение в гематологической и гистологической литературе.
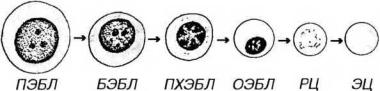
Процесс дифференцировки предшественников эритроцитов в зрелые форменные элементы включает (рис. 9-3):
(1)уменьшение размеров клетки;
(2)выработку и накопление гемоглобина в цитоплазме;
(3)постепенное снижение содержания и в конечном итоге утрату всех органелл;
(4)изменение окраски цитоплазмы от интенсивно базофильной (в связи с большим числом полирибосом) до оксифильной (обусловленной присутствием гемоглобина);
(5)снижение, а в дальнейшем (в конце стадии оксифильного эритробласта) - утрату способности к делению;
(6)конденсацию ядра и его последующее удаление из клетки.
Рис. 9-3. Эритропоэз. ПЭБЛ - проэритробласт, БЭБЛ - базофильный эритробласт, ПХЭБЛ - полихроматофильный эритробласт, ОЭБЛ - оксифильный (ортохроматофильный) эритробласт, РЦ - ретикулоцит, ЭЦ - эритроцит.
Проэритробласт - крупная клетка (диаметром около 18-22 мкм) с большим сферическим ядром, содержащим мелкодисперсный хроматин и дватри бледных ядрышка. Цитоплазма умеренно базофильна вследствие присутствия свободных рибосом. Клетка интенсивно пролиферирует, давая начало эритробластам. Последние развиваются в составе так называемых эритробластических островков.
Эритробластические островки - особые структурные комплексы в миелоидной ткани, обеспечивающие развитие эритробластов. Их центр образован телом макрофага, который своими многочисленными отростками охватывает окружающие его в один-два слоя эритробласты. По мере созревания эритробласты центробежно смещаются по длине отростков макрофага, удаляясь от его тела и отодвигаясь на периферию эритробластического островка, одновременно приближаясь к поверхности венозного синуса.
- 258 -
Базофилъный эритробласт - меньших размеров, чем проэритробласт (1216 мкм), с более мелким ядром, содержащим умеренно конденсированный хроматин и ядрышки. Цитоплазма резко базофильна благодаря высокому содержанию полисом, активно синтезирующих гемоглобин. Базофильные эритробласты активно делятся.
Полихроматофильный эритробласт характеризуется более мелкими размерами, чем батофильный эритробласт (10-12 мкм); его ядро более компактно, чем в базофильном эритробласте, глыбки хроматина в нем распределены в виде спиц колеса, ядрышко не выявляется. Цитоплазма окрашивается полихромно: она воспринимает как основные красители (вследствие наличия в ней многочисленных полисом), так и кислые (из-за накопления в ней оксифилыю окрашивающегося гемоглобина). Окраска цитоплазмы может быть либо диффузной и однородной, либо сочетать оксифилыше и базофильные участки. Скопления гемоглобина вокруг ядра часто имеют вид оксифильного перинуклеарного ободка. По мере накопления гемоглобина полисомы и другие органеллы редуцируются. Способность клетки к делению сохраняется.
Железо, необходимое для синтеза гема, поступает в цитоплазму эригробластов из двух источников: (1) непосредственно из крови (где оно связано с белком трансферрином) - путем транспорта, опосредованного рецепторами трансферрина на поверхности эригробластов; (2) из цитоплазмы макрофагов, контактирующих с эритробластами в эритробластических островках. Железо в составе ферритина (в комплексе с белком) выделяется на поверхность эригробластов в виде частиц диаметром 6 нм, которые связываются с их гликокаликсом и далее переносятся в их цитоплазму механизмом микропиноцитоза. Небольише скопления ферритина диаметром 0.1-0.3 мкм (сидеросомы) можно выявить в цитоплазме. Согласно расчетам, второй механизм переносит в 1000 раз больше железа, чем первый.
Оксифилъный (ортохроматофильный) эритробласт (нормобласт)
образуется путем дифференцировки из полихроматофильного эритробласта. По размерам он чуть крупнее эритроцита. Обладает оксифильно окрашенной цитоплазмой, богатой гемоглобином, в которой органеллы почти полностью отсутствуют. Ядро мелкое, компактное, пикнотическое, расположено эксцентрически. Способность к делению теряется.
Выталкивание ядра - наиболее важный этап в процессе превращения оксифильного эритробласта в ретикулоцит. Оно длится несколько минут и может происходить, когда клетка находится в составе
- 259 -
эритробластического островка или мигрирует через стенку кровеносных сосудов (синусов) костного мозга в кровоток. Этому процессу предшествует перестройка цитоскелета клетки, образующего структуру в виде манжетки, которая способствует активному выталкиванию ядра с тонким ободком окружающей его цитоплазмы за пределы клетки, где оно быстро фагоцитируется макрофагами.
Ретикулоцит представляет собой безъядерную (постклеточную) структуру. Его оксифильная цитоплазма, почти целиком заполненная гемоглобином, содержит остатки полирибосом и других органелл, которые выявляются при суправитальной окраске в виде базофильной сеточки. В кровотоке ретикулоцит в течение 24-48 ч превращается в зрелый эритроцит (см. главу 7).
Длительность всех этапов эритропоэза - от КОЕ-Э до образования зрелого эритроцита равна около 3-7 сут.
Регуляция процесса эритропоэза осуществляется рядом гуморальных факторов, из которых наибольшее значение имеют ИЛ-3 (стимулирует пролиферативную активность БОЕ-Э) и эритропоэтин (усиливает пролиферацию КОЕ-Э). Для нормального эритропоэза необходимы также железо, фолиевая кислота и витамин В12.
Эритропоэтин продуцируется у взрослого на 90% почкой, на 10% печенью (последняя, однако, служит главным его источником у плода) и вырабатывается в ответ на гипоксию. Его действие усиливается андрогенами, гормоном роста, тироксином и ослабляется эстрогенами (поэтому у женщин содержание эритроцитов и гемоглобина в крови ниже, чем у мужчин).
Недостаточная выработка эритропоэтина (например, при заболеваниях почек, некоторых эндокринных расстройствах - недостаточности гипофиза, щитовидной железы, коркового вещества надпочечника, мужском гипогонадизме) может вызывать развитие анемии, которая излечивается введением рекомбинантного эритропоэтина.
Применение эритропоэтина в качестве допинга у спортсменов для повышения физической работоспособности основано на увеличении переноса кислорода возросшим числом эритроцитов в крови. Последнее, однако, чревато риском развития тромботических осложнений из-за повышенной вязкости крови.
- 260 -