Материал: В-Л-Быков-Цитогогия_и_общая_гистология
фабрициевой сумке (Buisa Fabricii), с чем и связано название этих клеток. У млекопитающих, в том числе у человека, В-лимфоциты развиваются из клетокпредшественников в красном костном мозге (у плода - первоначально в желточном мешке, печени и селезенке). В ходе пролиферации и дифференцировки в костном мозге у этих клеток в отсутствие антигенной стимуляции происходит реаранжировка части генома, ответственной за антигенную специфичность В-лимфоцитов. В р-зультате образуются клетки с огромным (порядка 109 вариантов) репертуаром антиген-распознающих рецепторов.
Взаимодействие В-лимфоцитов с антигенами и их участие в иммунных реакциях:
1.Из костного мозга наивные В-лимфоциты попадают в кровь, неся на
своей поверхности молекулы иммуноглобулинов (IgM), которые представляют собой специфические антиген-распознающие рецепторы (до 104-105/клетку), а также характерные маркеры CD19, CD20, CD21, CD22 и CD23. В-лимфоциты экспрессируют на плазмолемме молекулы МНС I и II классов, рецепторы к СЗкомпоненту комплемента и Fcучасткам молекул иммуноглобулинов.
2.Из крови, в которой В-клетки составляют 10-2.0% циркулирующих лимфоцитов, они направляются в периферические иммунные органы и заселяют их В-зависимые зоны. В последних в результате взаимодействия с антигенами (и Тх) происходит их активация и пролиферация. Она завершается созреванием
идифференцировкой активированных В-лимфоцитов в плазматические клетки, продуцирующие антитела, и В-клетки памяти. Антитела вырабатываются и самими активированными В-лимфоцитами, однако их основным источником в организме служат плазматические клетки.
3.Часть активированных В-лимфоцитов с током лимфы возвращается в кровь, а из нее попадает в различные органы (в особенности, в очага воспаления), где они превращаются в плазматические клетки.
Роль В-лимфоцитов в местной иммунной системе слизистых оболочек. Значительная доля активированных В-лимфоцитов направляется в слизистые оболочки, в особенности, в те, которые послужили входными воротами для данного антигена. Слизистые оболочки всех систем организма содержат значительное количество лимфоидной ткани, которая обозначается термином MALT (аббревиатура англ. Mucosa-Associated Lymphoid Tissue -
ассоциированная со слизистыми обо-
- 236 -
лочками лимфоидная ткань) и образует основу местной иммунной системы слизистых оболочек. Здесь активированные В-лимфоциты дифференцируются в плазматические клетки, в том числе вырабатывающие секреторные IgA, которые обеспечивают местный гуморальный иммунитет слизистых оболочек, защищающий их от колонизации микроорганизмами и от возможного внедрения микробов в ткани.
Активация В-лимфоцитов происходит под влиянием двух сигналов:
1)антигена, связывающегося со специфическим иммуноглобулиновым рецептором на поверхности В-лимфоцита;
2)контактного взаимодействия В-лимфоцита с Тх, секретируюшим ряд интерлейкинов. Это взаимодействие Тх осуществляют лишь с собственными В- лимфоцитами, обладающими соответствующими детерминантами МНС, т.е. оно является МНС-рестриктированным. Необходимость во втором сигнале отсутствует при воздействии тимус-независимых антигенов (см. ниже).
Механизм активации В-лимфоцитов включает последовательность явлений, сходную с наблюдаемой при активации Т-лимфоцитов (см. выше) и завершается усилением экспрессии ряда генов, в частности, связанных с пролиферацией клеток и их дифференцировкой в плазматические клетки - антителопродуценты.
Взаимодействие В-лимфоцитов с антигенами протекает неодинаково и зависит от природы антигенов, что позволило разделить последние на тимусзависимые и тимус-независимые.
1.Тимус-зависимые антигены неспособны активировать В-лимфоциты в отсутствие второго сигнала, обусловленного Тх, что послужило основанием к их наименованию. К этой группе относится большинство существующих антигенов.
2.Тимус-независимые антигены могут эффективно стимулировать В- лимфоциты (вызывая их активацию с последующей пролиферацией и дифференцировкой) без участия второго сигнала. В эту группу входит лишь небольшое число антигенов с многократно повторяющимися эпитопами, которые перекрестно связывают мембранные иммуноглобулиновые рецепторы В-лимфоцитов. К таким антигенам относятся, например, высокомолекулярные полисахариды микроорганизмов.
Взаимодействие В-лимфоцитов с Тх
1. Антиген-распознающие рецепторы В-лимфоцита специфически связывают антиген, который далее поглощается механизмом рецептор-
- 237 -
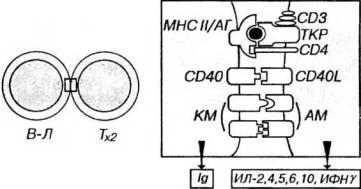
но-опосредованного эндоцитоза, подвергается процессингу и экспрессируется на поверхности В-лимфоцита в виде пептидов, связанных с молекулами МНС II класса (рис. 8-7).
2.Тх, ранее активированный данным антигеном в ходе взаимодействия с АПК, распознает комплекс молекулы МНС II класса/антиген на поверхности В- лимфонита. При этом в Тх происходит внутренняя перестройка, отчасти сходная с наблюдаемой в Тк при контакте с клеткой-мишеныо - ориентация его органелл в направлении В-клетки. Благодаря этому, очевидно, Тх способен осуществлять направленную секрецию интерлейкинов (главным образом, ИЛ-2, -4, -5, -6, -10) и ИФНγ на поверхность В-лимфоцита.
3.Указанные вещества активируют В-лимфоциты, стимулируют их пролиферацию и дифференцировку в плазматические клетки, способствуют переключению класса вырабатываемых антител. Активацию В-лимфоцита обеспечивают и ею непосредственные контактные взаимодействия с Тх, включающие связывание рецептора СD40 на плазмолемме В-лимфоцита с его лигандом (СD40L), экспрессируемым на поверхности активированною Тх. Взаимодействие СD40-СD40L необходимо для последующего переключения клетки с синтеза IgМ на продукцию иммуноглобулинов других изотопов IgG и IgA.
Рис. 8-7. Взаимодействие В-лимфоцита с Тх. Антиген (АГ) поглощается В- лимфоцитом, подвергается процессингу и экспрессируется на его поверхности в комплексе с молекулами МНС II класса (МНС II). Этот комплекс распознается Тх2 (посредством TKP и CD4), который секретирует ряд интерлейкинов на поверхность В- лимфоцита, активируя его, стимулируя пролиферацию и дифференцировку в плазматическую клетку, продуцирующую иммуноглобулины. Активации В-лимфоцита способствует контактное взаимодействие рецептора CD40 на его плазмолемме с лигандом CD40L на поверхности активированного Тх. Эффективность кооперации клеток повышается в результате взаимодействия костимулирующих молекул (КМ) на поверхности В-лимфоцита с адгезионными молекулами (AM) на плазмолемме Тх2.
- 238 -
Первичный гуморальный иммунный ответ развивается при первой встрече с антигеном и вызывает выработку небольшого количества антител. По прошествии определенного времени уровень антител обычно существенно падает.
Вторичный гуморальный иммунный ответ возникает при повторном попадании антигена и характеризуется быстрым развитием и продукцией больших количеств антител. Он обеспечивается благодаря активности В- и Т- лимфоцитов памяти (Вп и Тп). Эти клетки образуются при начальном воздействии антигена.
В-клетка памяти (Вп) - долгоживущая клетка, обеспечивающая быструю пролиферацию и дифференцировку В-лимфоцитов в плазмоциты при повторном контакте с антигенами. В отличие от виргильных лимфоцитов, которые в периферических иммунных органах живут всего несколько дней и погибают, если не встречаются со своим специфическим антигеном, Вп могут жить в течение нескольких месяцев и даже лет, не делясь и участвуя в рециркуляции.
ПЛАЗМАТИЧЕСКИЕ КЛЕТКИ
Плазматическая клетка (плазмоцит) - неподвижная или очень слабо подвижная, короткоживущая (2-3 сут, по другим сведениям - до 10-30 сут.) клетка - конечный этап развития В-лимфоцита. В ходе дифференцировки из В- лимфоцита она утрачивает рецепторы к СЗ-компоненту комплемента, мембранные иммуноглобулины и молекулы МНС, а также маркеры СD19 и
СD21.
Функциональные свойства плазматических клеток. Функция плазматических клеток заключается в обеспечении гуморального иммунитета путем выработки антител. За 1 секунду каждый плазмоцит синтезирует до нескольких тысяч молекул иммуноглобулинов (более 10 млн. молекул в час). Продуцируемые иммуноглобулины относятся к пяти классам (см. ниже), причем плазматические клетки способны переключаться с выработки иммуноглобулинов одного класса на другой.
Переключение классов продуцируемых иммуноглобулинов (переключение изотипов) происходит в развивающихся плазматических клетках примерно с 1- суточным интервалом - с IgM на IgG или IgА - без изменения их антигенсвязывающего участка. Процесс переключения связан с обратимыми изменениями процессинга транскриптов РНК. а также с необратимой рекомбинацией соответствующих участков ДНК. Одна клетка может синтезировать до трех классов иммуно-
- 239 -
глобулинов одновременно. Процесс переключения классов иммуноглобулинов контролируется цитокинами. Описаны иммунодефицитные состояния, связанные с нарушением переключения изотопа IgМ на IgG или (см. ниже).
Созревание аффинности продуцируемых иммуноглобулинов - выработка вновь образующимися плазмоцитами антител с возрастающей аффинностью (сродством) к данному антигену. Это явление служит отражением процессов соматических гипермутаций и селекции активированных В-лимфоцитов с высокоаффинными рецепторами, которые происходят в периферических органах кроветворения и иммуногенеза при продолжительном антигенном воздействии (см. главу 9).
Распределение плазматических клеток в организме. В норме плазматические клетки не циркулируют в крови. Они располагаются в красном костном мозге (составляя 1-2% его клеток), лимфатических узлах (преимущественно в мозговых тяжах и герминативных центрах), в белой пульпе селезенки. Значительное количество этих клеток характерно для рыхлой волокнистой соединительной ткани (см. главу 10), в особенности, образующей собственную пластинку слизистых оболочек (см. выше) и строму различных желез - например, слезных, слюнных, молочных (где преобладают клетки, продуцирующие IgA).
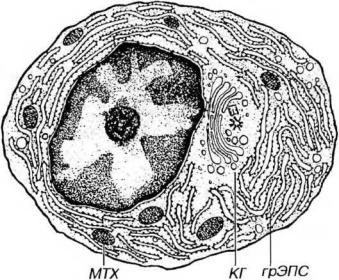
Рис. 8-8. Ультраструктурная организация плазматической клетки. КГ - комплекс Гольджи, МТХ - митохондрии.
- 240 -