Материал: В-Л-Быков-Цитогогия_и_общая_гистология
Взаимодействие Т-лимфоцитов с антигенами и их участие в иммунных реакциях
Т-лимфоциты способны взаимодействовать с антигеном (соответствующим по специфичности их ТКР) только, если он представлен им особыми АПК, в которых он ранее был подвергнут процессингу. В результате взаимодействия с антигеном Т-лимфоциты активируются, пролиферируют (экспансия клона), секретируют разнообразные цитокины (лимфокины), вновь поступают в кровь, а из нее - повторно в ткани (процесс рециркуляции), где они и осуществляют свои защитные функции. Часть лимфоцитов превращается в долгоживущие Т-клетки памяти (с фенотипом CD45RO+), которые сохраняют в течение всего времени своего существования усиленную экспрессию ТКР и ряда маркеров, что определяет их высокую чувствительность к повторному воздействию данного антигена. Рециркулирующие Т-лимфоциты живут до 4-6 мес., Т-клетки памяти сохраняются преимущественно в лимфоидных органах в течение многих лет. Специфика участия Т-лимфоцитов в различных защитных реакциях обусловлена их принадлежностью к одной из функциональных групп (субпопуляций).
Основные субпопуляции Т-лимфоцитов включают: Тх - Т-хелперы (от англ. help - помогать). Тс - Т-супрессоры (от англ. suppress - подавлять), Тк - Т- киллеры (Т-нитотокснческие лимфоциты), Тгзт - Т-клетки ГЗТ (гиперчувствительности замедленного типа) и Тп - Т- клетки памяти.
Функциональные группы Т-лимфоцитов объединяют 1) регуляторные клетки, влияющие на межклеточные взаимодействия (Тх и Тс), 2) эффекторные клетки, непосредственно осуществляющие защитные реакции (Тк и Тгзт), и 3) клетки памяти, сохраняющие иммунологическую "память" о первичном контакте с антигеном (Тп).
Активация Т-лимфоцитов требует распознавания ими, как минимум, двух сигналов:
1)эпитопа антигена в комплексе с молекулами МНС на плазмолемме АПК. Этот комплекс распознается с помощью ТКР и CD4или С08-компонента на мембране Т-лимфоцитов;
2)цитокинов или их комбинаций.
Рестрикция по МНС (от лат. restrictio - ограничение) - способность определенных субпопуляций Т-лимфоцитов взаимодействовать лишь с собственными клетками, экспрессирующими молекулы МНС, свойственные данному организму - важное условие процесса их активации.
- 226 -
Молекулы CD4 и CD8 служат дополнительными рецепторами (корецепторами) молекул МНС I и II классов, соответственно. Благодаря потребности в комбинированном активационном сигнале предотвращаются возможные реакции как на антиген, который не подвергся необходимому процессингу и представлению, так и на собственные антигены.
Адгезионные молекулы (лиганды) на поверхности Т-лимфоцитов путем своих взаимодействий с добавочными костимулирующими молекулами на поверхности АПК обеспечивают более эффективную активацию Т-лимфоцитов.
Нарушения взаимодействия комплекса МНС/антиген с ТКР, связанные с молекулярными дефектами экспрессии ТКР, дефицитом молекул МНС и коактивационных сигналов обусловливают развитие иммунодефицитных состояний различной тяжести.
Т-хелперы (Тх) играют основную роль в деятельности иммунной системы - распознавании антигена, запуске реакций клеточного и гуморального иммунитета, регуляции взаимодействий Т-лимфоцитов друг с другом и между Т- и В-лимфоцитами, продукции лимфокинов. Основная их функция - стимулирующее (хелперное) влияние на эффекторные клетки. Они экспрессируют на своей поверхности ТКР и молекулу CD4.
Распознавание антигенов и активация Тх. Распознавание комплекса молекулы МНС II класса/антиген, который находится на поверхности АПК (дендритной АПК, макрофага или В-лимфоцита), осуществляется посредством ТКР и CD4 на мембране Тх (рис. 8-2). АПК при этом выделяет ИЛ-1, воздействующий на Тх. Активация Тх, развивающаяся в результате этих сигналов, представляет собой закономерную последовательность, которая включает: (1) активацию фосфолипазы с образованием инозитолтрифосфата и диацилглицерола, (2) повышение уровня внутриклеточного кальция, (3) включение каскада внутриклеточных протеинкиназ, (4) усиление транскрипции генов, кодирующих продукты активации (цитокины и их рецепторы) и (5) пролиферацию соответствующего клона Тх.
Активированные Тх (а) экспрессируют ряд костимулирующих молекул (которые усиливают взаимодействия между клетками) и (б) выделяют лимфокины, регулирующие деятельность макрофагов, Т- и В-клеток.
- 227 -
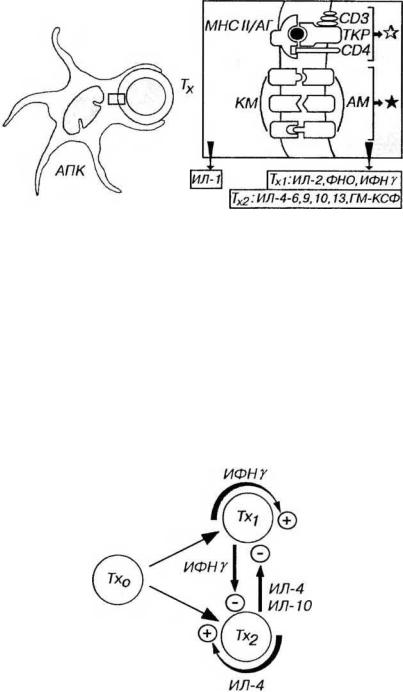
Рис. 8-2. Распознавание антигенов и активация Тх при его взаимодействии с АПК. Эпитоп антигена (АГ) в комплексе с молекулами МНС II класса (МНС II), который находится на поверхности дендритной АПК, распознается посредством ТКР и СD4 на мембране Тх, что обеспечивает главный активационный сигнал (обозначен светлой звездочкой). При этом АПК воздействует на Тх путем выделения ИЛ-1. Более эффективная активация Тх достигается добавочным сигналом (черная звездочка) в результате взаимодействия костимулирующих молекул (КМ) на поверхности АПК с адгезионными молекулами (АМ) на плазмолемме Т-лимфоцитов. При активации Тх выделяют ряд цитокинов, состав которых различается у Тх1, и Тх2.
Рис. 8-3. Развитие Тх и взаимодействие их подклассов. Тх1, и Тх2 развиваются из общего предшественника (Тх0), причем направление развития определяется многочисленными факторами (см. текст). Цитокины, выделяемые Тх каждого подкласса, усиливают образование клеток своего подкласса (стрелки со знаком "+") и угнетают развитие и активность клеток другого подкласса (стрелки со знаком "-").
- 228 -
Подклассы Тх (Тх1 и Тх2) различаются характером цитокинов (лимфокинов), секретируемых при активации, экспрессией некоторых костимулнрующих адгезивных молекул, а также функциональной ролью в различных звеньях иммунитета. Поддержание баланса между подклассами Тх определяет течение и исход инфекционных и аутоиммунных заболеваний (см. ниже). Оно осуществляется благодаря тому, что клетки каждого подкласса:
(1) усиливают дифференцировку "наивных” Т-клеток в данный (свой) подкласс Тх. Основными аутокринными факторами роста служат ИФНγ (для Тх1,) и ИЛ-4 (для Тх2).
(2) угнетают развитие и активность клеток другого подкласса (рис. 8-3). Так, ИФНγ (вырабатываемый Тх,), ингибирует образование Тх2, а ИЛ-4 и (или) ИЛ-10 (продуцируемые Тх2) - тормозят образование Тх,.
Тх1 отвечают преимущественно за реакции клеточного иммунитета и воспаления, частично - за некоторые реакции гуморального иммунитета (связанные с деятельностью фагоцитов). Они выделяют ИЛ-2, ИФНγ, ФНО и экспрессируют рецепторы к ИЛ-2. ИФНγ стимулирует, главным образом, макрофаги; ИЛ-2 усиливает пролиферацию Т-лимфоцитов (Тх и Тк), активирует NК-клетки, Т- и В-лимфоциты. Последние под влиянием ИФНγ переключаются на продукцию комплемент-связывающих и опсонизирующих антител (класса IgG), которые усиливают реакции фагоцитоза. ФНО и ИФНγ, привлекая и активируя гранулоциты, способствуют развитию воспалительных реакций.
Тх2 - стимулируют реакции гуморального иммунитета, участвуют в ряде регуляторных механизмов. Они выделяют ИЛ-4, -5, -6, -9, -10, -13, ГМ-КСФ. За счет действия указанных лимфокинов происходит активация В-лимфоцитов. усиливается их пролиферация и дифференцировка в плазматические клетки, вырабатывающие IgM, IgA, IgB (не связывающие комплемент), а также IgЕ (что стимулирует развитие ал¬ергических реакций). ИЛ-5 усиливает рост и активность эозинофилов, комбинация ИЛ-4 + ИЛ-10 - тучных клеток и базофилов, Регуляторная функция Тх2 связана с тем, что ряд секретируемых ими цитокинов обладает противовоспалительной активностью. Вследствие этого Тх2 угнетают реакции острого и хронического воспаления, включая ГЗТ. Во многих случаях это предотвращает нежелательное повреждение тканей, однако чрезмерная активность Тх2 связана с пониженным иммунитетом по отношению к внутриклеточным микроорганизмам.
Соотношение между Тх1 и Тх2 определяет характер течения и исход различных инфекций (вызванных вирусами, бактериями, гриба-
- 229 -
ми, простейшими и гельминтами), а также аллергических и аутоиммунных заболеваний. Так, при большинстве изученных инфекций человека преобладание Tx1 способствует высокой эффективности клеточных защитных реакций, а нарастание содержания Тх2 часто указывает на неблагоприятное течение заболевания.
Развитие Tx1 и Тх2 осуществляется из общего предшественника (Тх0), причем на его преимущественное направление (поляризацию) влияют многочисленные факторы: (1) концентрация и химическая природа антигена, (2) пути его введения, (3) костимулирующие адгезивные сигналы, полученных от АПК, (4) воздействие цитокинов, выделяемых рядом клеток микроокружения - макрофагами, лимфоцитами, тучными клетками, базофилами и эозинофилами,
(5) генетическая предрасположенность индивидуума.
Относительное содержание Тх в крови в норме у здорового человека составляет около 2/3 циркулирующих Т-лимфоцитов.
Снижение содержания Тх отмечается при различных врожденных и приобретенных иммунодефицитных состояниях. В частности, оно может быть результатом повреждающего действия ВИЧ (проникающего в эти клетки путем прикрепления к молекуле CD4) и характерным признаком СПИДа. Именно внутри Тх образуются более 99% из 1010 новых вирусных частиц, появляющихся в организме ВИЧ-инфицированного ежедневно. Причины возникновения дефицита Тх остается окончательно не выясненными; предполагаемые механизмы включают: 1) непосредственное разрушение Тх вирусом, 2) индукцию вирусом апоптоза Тх, 3) разрушение инфицированных Тх цитотоксическими лимфоцитами. Следствием потери значительной части Тх служит иммунная недостаточность, проявляющаяся в развитии тяжелых инфекций, в частности, вызываемых условно-патогенными микроорганизмами, которые практически безвредны для здорового человека, опухолей (например, саркомы Капоши), неврологических расстройств.
Т-супрессоры (Тс) распознают эпитопы антигенов в комбинации с продуктами системы МНС 1 класса. На их поверхности имеются ТКР и молекулы CD8. Клетками-мишенями Тс являются В-лимфоциты, Тх и Тк (см. ниже). Функция Тс заключается в угнетении активности иммунных реакций путем непосредственного контактного воздействия на указанные клетки или секреции угнетающих (супрессорных) факторов. Тс подавляют развитие аутоиммунных реакций. В последние годы высказывается мнение, согласно которому роль Тс могут играть Тх1 и Тх2, которые вырабатывают ряд цитокинов, способных угнетать активность лимфоцитов и макрофагов.
- 230 -