Материал: Полимеразная цепная реакция
2)Высокая специфичность (т.е. низкая вероятность ложноположительного результата).
3)Относительная быстрота.
4)Пригодность для всех видов возбудителей, в том числе для некультивируемых, которых нельзя выявить иными методами.
Механизм амплификации при проведении ПЦР во многом повторяет механизм репликации ДНК, происходящий в клетке.
СТРОЕНИЕ ДНК И РНК
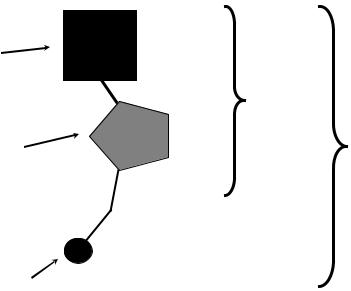
Дезоксирибонуклеиновая кислота (ДНК) представляет собой линейный полимер, состоящий из остатков нуклеотидов. Нуклеотиды представляют собой фосфорные эфиры нуклеозидов, которые, в свою очередь, состоят из дезоксирибозы и азотистого основания (рис. 1).
АЗОТИСТОЕ ОСНОВАНИЕ
ДЕЗОКСИ- |
НУКЛЕОЗИД |
НУКЛЕОТИД |
РИБОЗА |
|
|
5’ |
3’ |
|
|
|
ФОСФОРНАЯ
КИСЛОТА
Рис. 1. Строение нуклеотида.
Азотистые основания в молекуле ДНК бывают 4-х видов: пуриновые (аденин и гуанин) и пиримидиновые (цитозин и тимин). Нуклеотиды в составе молекулы ДНК соединены друг с другом с помощью остатков фосфорной кислоты по принципу «голова к хвосту». Фосфорная кислота связывает 3-й углеродный атом дезоксирибозы одного нуклеотида и 5-й атом углерода следующего нуклеотида. Один из концевых нуклеотидов имеет свободный 3-й атом углерода дезоксирибозы, и этот конец цепи ДНК обозначается как 3'-конец. Другой конец цепи ДНК имеет свободный 5-й углерод дезоксирибозы и обозначается, соответственно, как 5'-конец.
Гетероциклические основания в составе ДНК могут связываться друг с другом при помощи водородных связей. При этом аденин связывается только с тимином 2 водородными связями, а гуанин – с цитозином при помощи 3 водородных связей.
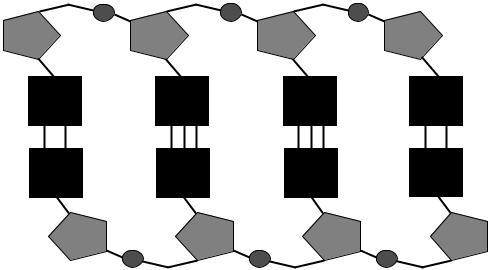
Молекула ДНК обычно состоит из 2 полинуклеотидных цепей, которые удерживаются вместе при помощи водородных связей между гетероциклическими основаниями. Соответственно, основания входящие в состав цепей ДНК, должны соответствовать друг другу: напротив аденина одной цепи должен находиться тимин другой цепи, а напротив гуанина – цитозин. Таким образом, цепочки ДНК должны быть комплементарны друг другу и, зная нуклеотидную последовательность одной из цепей, легко узнать нуклеотидную последовательность комплементарной цепи. Цепи в составе молекулы ДНК расположены антипараллельно друг другу, т.е. напротив 5’-конца одной цепи находится 3’-конец комплементарной цепи (рис. 2).
3’ |
5’ |
|
A |
G |
С |
T |
А |
аденин |
|
|
|
|
||
|
|
|
|
G |
гуанин |
|
|
|
|
С |
цитозин |
T |
C |
G |
А |
Т |
тимин |
|
|
||||
5’ |
|
|
|
3’ |
|
Рис. 2. Строение двухцепочеченой ДНК (пояснение в тексте).
Гетероциклические основания ДНК гидрофобны, поэтому в водной фазе ДНК образует спиралевидную вторичную структуру, уменьшая их контакт с водой. Выделяют 3 основных формы вторичной структуры ДНК (А-, В- и Z-). Самой распространенной и известной является В-форма, описанная Дж. Уотсоном и Ф. Криком в 1953 г. В-форма представляет собой спираль, в которой обе цепи ДНК закручены друг относительно друга и вокруг общей оси. Витки спирали одинаковы, и на каждый приходятся 10 нуклеотидных остатков, при этом гетероциклические основания обращены внутрь спирали, а углеводные остатки – наружу.
РНК также представляет собой линейный полимер из остатков нуклеотидов, связанных между собой остатками фосфорной кислоты. Нуклеотиды в составе РНК вместо дезоксирибозы содержат рибозу и вместо тимина – урацил. Молекулы РНК обычно являются одноцепочечными, лишь у некоторых вирусов (например, у ротавирусов) есть двухцепочечные РНК. Одноцепочечные РНК не
образуют двойных спиралей, но в составе молекулы могут встречаться комплементарные участки, которые самопроизвольно образуют связи между собой. В результате РНК укладывается во вторичную структуру, в составе которой присутствуют петли. Основание этих петель образовано комплементарными участками РНК, связанными между собой с помощью водородных связей. РНК менее стабильна, чем ДНК, и легче разрушается под воздействием внешних факторов.
В клетке РНК является «посредником» в реализации генетической информации, содержащейся в ДНК. Однако у многих вирусов РНК является основным носителем генетической информации.
РЕПЛИКАЦИЯ И ТРАНСКРИПЦИЯ НУКЛЕИНОВЫХ КИСЛОТ В КЛЕТКЕ. ОБРАТНАЯ ТРАНСКРИПЦИЯ.
ДНК является основным носителем генетической информации всех клеточных форм, поэтому цикл деления любой клетки обязательно включает в себя стадию репликации, т.е. удвоения содержащейся в ней ДНК. Цепи в составе молекулы ДНК комплементарны друг другу, и репликация ДНК происходит полуконсервативно, т.е. происходит разъединение цепей в молекуле ДНК, после чего к каждой из получившихся матричных цепей достраивается комплементарная. В результате образуются две идентичные молекулы ДНК.
Репликация РНК (т.е. синтез РНК на матрице РНК) в клетке в норме не происходит. Синтез РНК в клетке (транскрипция) осуществляется на матрице геномной ДНК.
Цепь ДНК, с которой происходит транскрипция, является кодирующей и обозначается как минус-цепь (–ДНК), комплементарная ей цепь является некодирующей и обозначается как плюс-цепь (+ДНК). При репликации ДНК на матрице плюс-цепи ДНК синтезируется комплементарная ей минус-цепь и наоборот.
Схема репликации ДНК лучше всего изучена на примере бактерии E. coli. Этот процесс включает в себя 4 стадии:
1)Разъединение цепей ДНК
2)Синтез РНК-затравок
3)Достройка комплементарных цепей ДНК
4)Удаление РНК-затравок
Врепликации участвуют следующие ферменты:
1)Хеликаза, разделяющая нити ДНК
2)Праймаза, формирующая РНК-затравки для синтеза новых цепей ДНК.
3)ДНК-полимераза. Это фермент, который осуществляет синтез ДНК. Выделяют несколько разновидностей ДНК-полимераз, в репликации ДНК ключевую роль играют ДНК-полимераза I и III. ДНК-полимераза III синтезирует цепь комплементарную цепь, тогда как ДНК-полимераза I удаляет из цепи РНК-затравки.
Набор ферментов, участвующих в репликации ДНК, может иметь некоторые различия у разных групп живых существ, в том числе у разных групп бактерий, но общая схема процесса остается одинаковой.
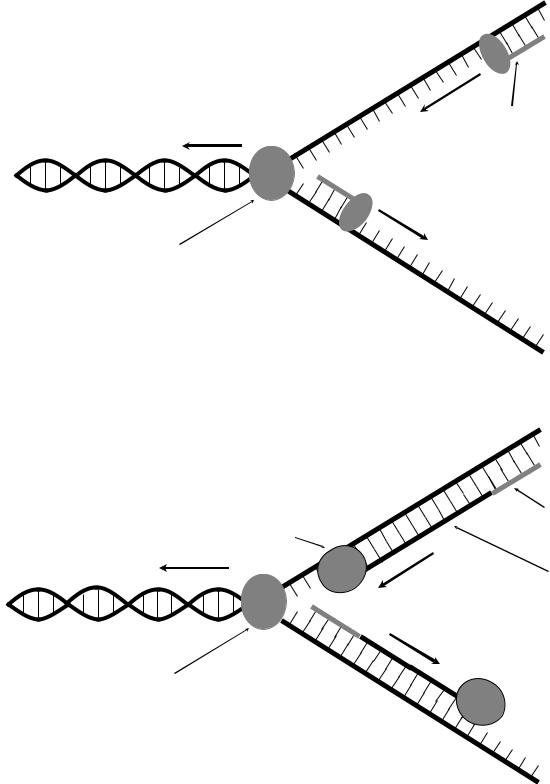
Молекулы ДНК состоят из 2 цепочек, которые перед началом репликации необходимо разъединить. Эту функцию выполняет фермент хеликаза, формирующий движущуюся репликативную вилку, которая состоит из двух разъединенных цепей и оставшейся неразъединенной части ДНК. К разъединенным цепям ДНК присоединяется SSB-белок, который препятствует их обратному соединению.
К каждой из образовавшихся матричных цепей ДНК необходимо с помощью ДНК-полимеразы достроить комплементарную цепь, получив, таким образом, 2 идентичные молекулы ДНК. Однако ДНК-полимеразы не способны синтезировать комплементарную цепь «с нуля», а могут лишь удлинить имеющуюся, поэтому синтез новых цепей ДНК невозможен без затравки. Фермент праймаза синтезирует на обеих матричных цепях ДНК короткие цепочки РНК длиной несколько нуклеотидов (рис. 3А). Используя эти цепочки РНК как затравки, ДНК-полимеразы III «удлиняют» их, достраивая комплементарные цепи ДНК. Движение ДНК-полимеразы по матричной цепи ДНК идет строго в одном направлении: от 3'-конца к 5'-концу. На одной из матричных цепей ДНКполимераза III движется по ходу расплетания ДНК (т.е. по ходу движения хеликазы). Но так как цепи в молекуле ДНК антипараллельны друг другу, то на второй цепи движение ДНК-полимеразы III осуществляется против направления расплетания цепей. В этом случае комплементарная цепь синтезируется короткими фрагментами (фрагментами Оказаки): по мере расплетания цепи праймаза синтезирует РНК-затравки, после чего ДНК-полимераза III «заполняет» промежутки между ними (рис. 3Б).
После завершения репликации образуются 2 идентичные молекулы ДНК, каждая из которых содержит одну матричную и одну вновь синтезированную цепь. РНК-затравки, оставшиеся во вновь синтезированных цепях ДНК, удаляются с помощью фермента ДНК-полимераза I. Эта разновидность ДНКполимеразы способна не только синтезировать ДНК, но также обладает экзонуклеазной активностью: двигаясь по матричной цепи, она способна отщеплять находящиеся перед ней нуклеотиды. ДНК-полимераза I разрушает РНК-затравки, одновременно заполняя образующиеся бреши ДНК.
3’-конец
ПРАЙМАЗА 
РНК-ЗАТРАВКА
ХЕЛИКАЗА
А
5’-конец
3’-конец
РНК-
ЗАТРАВКА
ДНК-ПОЛИМЕРАЗА III
СИНТЕЗИРУЕМАЯ ЦЕПЬ ДНК
ХЕЛИКАЗА
Б
5’-конец
Рис. 3. Схема репликации ДНК in vivo. А: разъединение цепей ДНК с участием хеликазы и синтез РНК-затравок с участием праймазы. Б: достройка комплементарных цепей ДНК сДНК-полимеразы III. Стрелками указано направление движения ферментов.