Материал: Мхи Сахалина
Вот и выясняется, что ни та ни другая группа гипотез не решают вопроса о происхождении мохообразных достаточно убедительно. Гипотезы псилофитного происхождения привлекают больше сторонников и более распространены, чем водорослевые. Однако это не является следствием большей убедительности или доказанности их. Предпочтение какой-либо гипотезы сегодня не вывод из непреложных фактов, а личная точка зрения исследователя, не столько вытекающая из фактов, сколько обусловленная не всегда понятными «поворотами мысли». К тому же в некоторых из последних вариантов, исходящих из предположения, что мохообразные произошли от каких-то сравнительно просто устроенных, первичных гипотетических псилофитов, эти гипотезы в какой-то мере смыкаются с водорослевыми гипотезами. Ведь эти гипотетические первичные псилофиты в сущности и есть то промежуточное звено между водорослями и мохообразными, те полуводоросли-полумохообразные, о которых говорилось выше.
Таким образом, утверждать, что вопрос о происхождении мохообразных решен сколько-нибудь удовлетворительно, будет, пожалуй, рановато. Слишком много еще неясного, неопределенного, в том числе и в вопросах филогении этой группы. Да и трудно было бы ожидать иного: происхождение и филогения тесно между собою связаны, и нерешенность одной проблемы сразу же ставит массу вопросов в другой.
Почти без преувеличений можно сказать, что в филогении мохообразных нет пока убедительных решений основных вопросов. Так, мы не знаем, в какой последовательности появлялись классы и подклассы мохообразных. Более или менее единодушны бриологи в признании за печеночниками большего возраста, чем за листостебельными мхами. Предположим, это - верная точка зрения. Но и она - скорее предположение, чем вывод из фактов. Совершенно неясным остается время появления антоцеротовых. Мы уже упомянули, что в ископаемом состоянии они неизвестны. Правильнее будет сказать, что неизвестны макроостатки. Споры обнаружены в отложениях третичного возраста. А ведь по многим признакам, например наличию хлоропласта с пиреноидом, именно антоцеротовые наиболее близко стоят к водорослям. Какова последовательность появления порядков, каковы их родственные связи? Ответа, четкого, доказательного, нет.
В попытках выйти из этих сложностей родилась точка зрения, исходящая из предположения, что мохообразные представляют собою полифилетическую по происхождению (возникшую от разных предков) группу, что одни предки их дали талломные формы, а другие - листостебельные. Те и другие, следовательно, развивались сами по себе, не превращаясь одна в другую. Бесспорно, такой подход снимает массу вопросов, касающихся истории мохообразных. Но возникают другие, и вопрос о непосредственных предках мохообразных почти совсем теряет ясность.
В оправдание бриологам надо сказать, что ответов на перечисленные вопросы нет не по их вине. Дело в объективных, очень больших трудностях. Ведь мохообразные представлены рядом далеко разошедшихся, часто очень слабо между собой связанных, мало родственных линий развития. Эти линии - циклы форм различного ранга от рода до класса, как бы замкнуты внутри себя, не связаны переходами друг с другом. Часть линий развития представлена крайне олиготипными или даже монотипными таксонами, нередко довольно высокого ранга. Например, монотипный порядок Schistostegales с одним видом Schistostega pennata, олиготипные порядки Takakiales с одним родом и двумя видами, Tetraphidales с двумя родами и тремя видами, Phyllothalliales с одним родом и двумя видами и т. п.
Такой характер родственных связей крайне затрудняет установление степени родства между отдельными группами мохообразных и построение системы бриофитов. Он свидетельствует об очень большой древности мохообразных, о вымирании многих промежуточных звеньев, когда-то связывавших ныне далеко друг от друга отстоящие группы.
И все это вместе очень затрудняет прояснение филогении мохообразных. К многочисленным трудностям надо добавить еще одну, сугубо бриологическую. В жизненном цикле мохообразных присутствуют две ядерных фазы - гаметофаза и спорофаза. И хотя одна без другой (именно спорофаза без гаметофазы, но не наоборот) жить не может, эволюция той и другой может идти и независимо, сама по себе, с разной скоростью, да подчас еще и в разных направлениях! Случаи, когда мы имеем относительно развитый, продвинутый гаметофит и примитивный или деградировавший спорофит, или прямо противоположные, с развитым спорофитом и примитивным или редуцированным гаметофитом нередки среди мохообразных. Примером первого может служить порядок Polytrichales, у представителей его высокоразвитый гаметофит с проводящей системой наиболее сложного для мохообразных типа и вместе с тем довольно примитивно устроенный спорофит с мелкими и малоподвижными зубцами перистома. Аналогичный пример показывают и сфагны: сложнейше устроенный гаметофит и весьма примитивный, во всяком случае, явно отставший от него в развитии спорофит. Противоположный пример - представители порядка Buxbaumiales: у них очень сложно устроенный крупный спорофит и мелкий, деградировавший (или примитивный?) гаметофит. Сложно и разнообразно устроены спорофиты у представителей семейства Splachnaceae при относительной простоте и однообразии в строении гаметофита.
Но всем этим трудности еще не исчерпываются. Добавляет их мозаичность в распределении признаков: какой-нибудь таксон, по всей сумме признаков, несомненно, принадлежащий определенному классу, вдруг обнаруживает признаки, свойственные другому классу, причем не всегда самому близкому. Так, колоколообразное строение спорового мешка наблюдается у антоцеротовых, а затем у сфагнов и андреевых мхов, но отсутствует у печеночников. Между тем в целом печеночники, несомненно, гораздо ближе к антоцеротовым, чем сфагновые и андреевые. А в свою очередь сфагны и андреевые, при всем их несомненном своеобразии, все же гораздо ближе к бриевым мхам, нежели к печеночникам и тем более к антоцеротовым. Посредством створок вскрываются коробочки у антоцеротовых, у многих печеночников и у андреевых. По всем остальным признакам и общей их сумме андреевые мхи ближе к другим подклассам листостебельных мхов, чем к печеночникам и антоцеротовым.
Однако бриологи не падают духом. И
продолжают пробиваться сквозь многочисленные трудности к познанию происхождения
и филогении мохообразных.
2.2 Как
устроены мохообразные
Мохообразные - небольшие многолетние или реже однолетние растения, талломные (слоевищные) или листостебельные, размножающиеся спорами. Многолетние мохообразные - вечнозеленые: таллом на зиму не отмирает, листья не опадают, и то и другое после периода зимнего покоя весной возобновляет жизнедеятельность. Период же зимнего покоя короче, чем можно было бы думать. Установлено, что многие мхи активно фотосинтезируют под снежным покровом.
Зеленое тело мохообразных, как
говорилось выше, гаметофит или гаметофаза. Оно может быть талломным или листостебельным.
Таллом - это просто лепешка или лента шириной в несколько миллиметров
(максимально до 5 см у южноамериканско-новозеландского печеночника Monoclea
forsteri)
и длиной в несколько сантиметров, чаще не более 4-6. Максимальную для талломных
печеночников длину - до 20 см - имеет все та же Monoclea
forsteri.
Таллом дихотомически ветвится, что особенно хорошо заметно, когда он имеет
форму неширокой ленты. Он всегда стелется и более или менее прижат к субстрату.
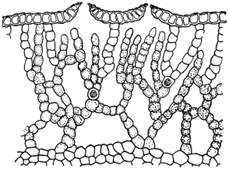
Рис. 1. Поперечный срез слоевища печеночника, относящегося к подклассу Маршанциевых, - Mannia brachypoda. Видны воздушные камеры, ассимиляционные нити (не достигающие верхнего слоя клеток клеточные ряды) и продушины в верхнем слое клеток слоевища. Пример довольно сложно устроенного слоевища.
Внутренне строение таллома довольно разнообразно. У одних видов он большей частью состоит из вполне однородных клеток. У других же печеночников из подкласса маршанциевых таллом устроен сложно (рис. 1). Верхняя стенка его образована одним слоем прозрачных клеток. В стенке имеются особые отверстия - продушины или норы, называемые иногда устьицами (но ничего общего с настоящими устьицами не имеющие). Эти продушины ведут в разделенные вертикальными перегородками и заполненные воздухом полости (камеры). Со дна их поднимаются нити, состоящие из хлорофиллоносных клеток. Эти нити называются обычно ассимиляционными или ассимиляторами.
Листостебельное растение мохообразного имеет ряд отличий от остальных листостебельных высших растений.
У всех без исключения бриофитов листья всегда сидячие, не имеют черешка. Пластинка листа прикрепляется непосредственно к стеблю. Она может располагаться перпендикулярно стеблю (подавляющее большинство листостебельных мхов), косо (большинство листостебельных печеночников) и продольно (только один вид из класса листостебельных мхов - Schistostega pennata, относящийся к отдельному монотипному порядку Schistostegales).
Лист устроен довольно разнообразно. Чаще всего он представляет собой однослойную (из одного слоя клеток) пластинку с жилкой или без нее. Форма - от линейной до округлой или даже поперечно-широкой (с шириной, превышающей длину). У листостебельных мхов пластинка листа всегда цельная, у печеночников часто в той или иной степени раздельная. Нередко лист продольно складчатый или поперечно-волнистый.
Поверхность листьев не всегда гладкая. Во многих случаях она «всхолмлена» выступающими частями клеток. В особенности разнообразны по форме, размерам, количеству (на одну клетку) выступы клеточной оболочки - папиллы.
Располагаются листья по стеблю большей частью спирально, и стебель чаще всесторонне облиственный.
У листостебельных печеночников (стебель у них обычно не бывает прямостоячим, в большинстве случаев он вытянут вдоль субстрата) листья располагаются двумя четко выраженными рядами, идущими по бокам стебля. У большинства видов есть, кроме того, и третий ряд листьев, расположенный внизу, на брюшной (обращенной к субстрату) стороне стебля. По форме и размерам эти брюшные листья обычно отличаются от спинных и носят особое название - амфигастрии.
У всех без исключения мохообразных отсутствуют корни. Прикрепление к субстрату (отчасти также поглощение влаги из почвенных растворов) осуществляется с помощью ризоидов. Они представляют собою нити из одного ряда клеток, разделенных косыми перегородками. Это у листостебельных мхов. У печеночников и антоцеротовых ризоиды всего лишь очень сильно вытянутые в длину одиночные клетки.
На гаметофите образуются гаметангии - антеридии и архегонии. И те и другие в принципе устроены одинаково у всех архегониатных растений. Антеридий - удлиненное, несколько мешковидное, у печеночников чаще округлое тело. Он имеет стенку из одного слоя стерильных клеток. Все внутреннее пространство его заполнено материнскими клетками антерозоидов - мужских гамет. При созревании антеридия на его удаленном от места прикрепления конце возникает отверстие, и снабженные каждый двумя жгутиками антерозоиды выходят наружу. Они способны самостоятельно двигаться в водной среде.
Архегоний - чаще узкоколбовидное или бутылковидное тело с узкой и длинной верхней частью (шейкой) и расширенной нижней (брюшком). Шейка имеет стенку из одного слоя клеток, брюшко - из нескольких. В брюшке располагается неподвижная яйцеклетка - женская гамета.
В результате оплодотворения (возможного только при наличии воды, только в ней могут двигаться антерозоиды) возникает зигота, дающая начало спорофиту или спорофазе. Наиболее часто, впрочем, его называют третьим именем - спорогоном. Спорофит - спороносный орган и состоит из трех частей: стопы, ножки и коробочки. Стопа - нижняя часть спорогона, с ее помощью последний внедряется в ткань гаметофита, прикрепляется к нему механически и получает от него питательные вещества и воду. Ножка - это стержень (у листостебельных мхов довольно прочный), на верхушке которого сидит коробочка. Коробочка - собственно и есть спорангий, орган, в котором образуются споры. У листостебельных мхов она представляет собою чаще бокаловидное образование с широким отверстием наверху (рис. 2), закрытым особым щитком с весьма обыденным названием - крышечка. Представьте себе туристский котелок с суженным основанием, закрытый крышкой. Это и будет коробочка наиболее распространенного у листостебельных мхов типа. Только для полноты картины котелок надо еще прикрыть сверху шапкой. Она будет изображать колпачок - специальное образование, закрывающее сверху коробочку. Форма и размеры колпачка очень разнообразны (рис. 3). Наиболее часто колпачок напоминает капюшон, разрезанный с одной стороны. В этом случае он называется клобуковидным. Но нередко колпачок имеет шапочковидную или колоколовидную форму. Обычно он голый, но довольно часто покрыт (редкими или густыми) волосками. В особенности характерно наличие густого покрова из волосков для представителей порядка, к которому относится хорошо известный мох - кукушкин лен, или политрихум обыкновенный (Polytrichum commune). По этому признаку и порядок и род получили название Polytrichum (в переводе с латинского языка означает многовласник).
В молодом возрасте коробочка, как правило, целиком или почти целиком закрыта колпачком, в зрелом - лишь частично. Роль и функции колпачка ясны - он защищает молодой спорогон и созревающие в нем споры от неблагоприятных воздействий внешней среды, главным образом резких колебаний температуры и влажности.

Рис. 2. Открытая коробочка
листостебельного мха. Хорошо виден простой перистом по краю устья коробочки.
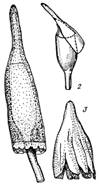
Рис. 3. Различные типы колпачков.
- колокольчатый представителя
порядка Encalyptales; 2 -
клобуковидный;3 - шапочковидный.
Удивляет не функциональное назначение колпачка, а его происхождение. Если весь спорогон - стопа, ножка, коробочка с крышечкой - возникли из зиготы, то колпачок - из стенки архегония. И, значит, он - не спорофит, а часть гаметофита! Странно, не правда ли? Но еще более странен колпачок некоторых печеночников - он возникнет не только из стенки архегония, но и из участков стебля или слоевища (таллома), частично срастающихся со стопкой архегония.
В отличие от крышки, закрывающей котелок, крышечка на созревающей коробочке листостебельных мхов лежит не свободно, а как бы прирастает к краю коробочки и составляет с нею одно целое. Между нижним краем крышечки и верхним краем коробочки (он называется устьем) располагаются один или несколько рядов особым образом устроенных гигроскопических клеток. Это колечко, клетки которого имеют резко утолщенные с одной стороны стенки. При созревании коробочки происходит ее частичное обезвоживание, подсыхание. Клетки колечка теряют воду. В них создается напряжение, так как, подсыхая, они уменьшаются в объеме, и неутолщенные части стенок начинают втягиваться внутрь. Когда напряжение и силы сцепления между клетками колечка и краями крышечки и коробочки становятся почти равными, тогда достаточно очень небольшого усилия и произойдет разрыв колечка. Оно распадается на отдельные клетки или куски из нескольких клеток. Крышечка при этом отпадает, отделяется от коробочки, и созревшие споры начинают высыпаться наружу. Усилие, которое разорвет колечко и обеспечит открывание коробочки, могут создать, например, легкое дуновение ветерка или насекомое, севшее на коробочку.
Высыпающиеся споры служат для бесполого размножения. Возникают они из особых клеток, называющихся материнскими. При образовании спор происходит редукционное деление, и они имеют уже гаплоидный набор хромосом. Споровый мешок, или археспорий, из которого образуются споры, не заполняет обычно всего внутреннего пространства коробочки. У очень многих видов листостебельных мхов и у антоцеротовых в центре коробочки располагается столбик-стерженек из стерильных клеток. Называется этот стерженек колонкой. Основная функция колонки - обеспечение споровой массы влагой и питательными веществами. Споровый мешок либо прикрывает колонку сверху наподобие шапки или колокола (у антоцеротовых, у сфагнов и у андреевых мхов), либо окружает ее в виде цилиндра, открытого и снизу и сверху (у бриевых мхов). Отсутствует колонка у печеночников.