Материал: Мейобентос зарослей макрофитов прибрежной зоны Новороссийской бухты
Мейобентос по глубинам распределен неравномерно.
Как правило, наибольшие количественные характеристики приурочены к
псевдолиторали и верхней сублиторали, и прежде всего к зоне фиталя. Эта же зона
наиболее богата по таксономическому составу фауны [Колесникова, 1991].
.5 Трофические связи в сообществе мейобентоса
Несмотря на малые размеры, представители мейобентоса обладают очень большой численностью. Высокая плотность поселений - одна из важных их особенностей. Достигая численности свыше 5 × 106 экз./м2, они обладают высокой плодовитостью, в основном коротким жизненным циклом, быстрым темпом роста, высокой интенсивностью метаболизма[Воробьева, 1999]. Организмы из состава мейобентоса играют важную роль в поддержании баланса органических веществ в донных осадках, т. к. через них проходит значительная доля всего энергопотока [Мокиевский, Малых, 2002].
На ухудшение условий существования мейобентос отвечает сокращением одних групп, доминированием двух - трех групп и повышением плотности организмов с минимальными размерами тела, обладающими, как правило, короткими циклами развития [Мейобентос в санитарно-биологическом аспекте, 1995].
В процессе жизнедеятельности представители мейобентоса, кроме детрита и диатомовых водорослей, потребляют значительное количество бактериальной и инфузориальной продукции. Мейобентос также в значительной мере существует за счет жизнедеятельности зоопланктона и макрозообентоса [Маккавеева, 1967].
Представители микро-, мейо- и макрозообентоса образуют сложные трофические связи, включающие питание не только мертвым органическим веществом и продуктами жизнедеятельности гидробионтов из состава других комплексов, но и живым органическим веществом.
Во всех сообществах мейобентосных животных
непрерывно осуществляется сложный механизм перестроек, обусловленный сезонными
изменениями в количественных и качественных характеристиках представителей
самого мейобентоса, их пищевых компонентов, макрозообентоса и ихтиофауны
[Колесникова, Мазгумян, 2002].
.6 Таксономический состав мейобентоса. Основные
группы мейобентосных организмов
В состав мейобентоса входят представители практических всех известных типов беспозвоночных, но относительное число микроскопических видов в разных таксонах различно.
Некоторые типы представлены исключительно видами мейобентосных размеров, в большинстве же типов известно лишь небольшое число мелких видов.
Степень изученности биологии различных мейобентосных групп варьирует в широких пределах.
Наиболее подробно изучены самые массовые группы - нематоды, гарпактициды, остракоды.
Большое число исследований посвящено таксономии, строению и образу жизни этих организмов. Сведения по биологии большинства редких и малочисленных групп скудны и отрывочны.
Для некоторых таксонов известен лишь факт
нахождения микроскопических представителей, сопровождающийся иногда кратким
очерком анатомии и морфологии [Воробьева, 1999]. Основные виды представителей
мейобентоса представлены в таблице 1.
Таблица 1 - Основные представители мейобентоса (Е. Б. Макавеева [1967])
|
Отряд |
Виды |
|
Coetenterata |
Agtaopheniapluma Plumutanalincoi Lucemaria campanula |
|
Polychaeta |
Fabriciasabelta Grubealimbata Harmothoe reticulate Nereiszonata Platynereisdumerilii Spirorbismolitaris Spirorbispusilta |
|
Loricata |
Acanthohitonafascicularis Lepidochitonacinerea |
|
Gastropoda |
Gibbulaadriatica Nana donovani Nana neritea Rissoasplendida Rissoamembranacea |
|
Bivalvia |
Mytilasterlieatus Mytilusgallaprovincialis |
|
Cirripedia |
Balanusimprovisus |
|
Cumacea |
Cumacea sp. |
|
Tanaidacea |
Lepcocheliasavignyi |
|
Isopoda |
Idoteaostroumovi Sphaeromaserratum Synisomacapito |
|
Amphipoda |
Apherusabispinosa Biancolinacuniculus Hyaledolffusi Hyaleperieri Hyalepontica Stenothoemonoculoides |
|
Decapoda |
Decapod asp. Decapodazoea Hippolitelongirostris |
|
Chironomida |
Chironomidae larvae |
|
Bryozoa |
Lepraliapallasiana |
|
Tunicata |
Diplosomalisterianum |
По суммарному числу видов мейобентос ненамного уступает макроскопическим беспозвоночным. Приблизительный расчет на основании списков видов для морей показывает, что число известных видов мейобентоса составляет около 25% от видового разнообразия бентоса. Доля мейобентоса от общего числа видов свободноживущих беспозвоночных в наиболее хорошо изученных морях (Белое, Баренцево) равна 16-20 %, но падает до 5-6 % в морях Сибири, что связано со слабой изученностью мейофауны. Локальное разнообразие мейофауны может быть сопоставимо и даже превышать разнообразие макробентосных видов [Мокиевский, 2009].
Чутко реагируя на изменения условий обитания, мейобентос в последние десятилетия также претерпел изменения в своем составе, численности и биомассе. Вероятно, эти перемены должны были отразиться на биологии связанных с мейобентосом организмов, в том числе и на естественном воспроизводстве донных рыб [Заика, Киселев, Михайлова, 1992].
Обилие и фаунистическое разнообразие эпифитона больше всего зависит от вида водоросли, на которой обитают животные в биотопе. Основными зарослеобразующими макрофитами Новороссийской бухты являются Cystoseira barbata и Cystoseira crinita. Таксономический и количественный сосав мейобентоса определялся в зарослях Cystoseirabarbata [Колесникова, 1981].
Заросли цистозиры, произрастая на прибрежной полосе скал и валунов, занимают значительные площади шельфа северо-восточной части Черного моря. Они служат субстратом, пищей и убежищем для многочисленных беспозвоночных, в том числе мейобентоса [Миловидова1969]. Известно, что обилие и фаунистическое разнообразие мейоэпифитона больше всего зависят от вида водоросли, на которой обитают животные, а также от степени прибойности и интенсивности водообмена в биотопе. Поэтому зарослевые биоценозы открытого прибрежья, по сравнению с защищенными участками моря, имеют ряд особенностей. На макрофитах, растущих в открытых незащищенных от волнового воздействия местах прибрежья, показатели численности мейобентоса могут быть ниже на порядок, чем на тех же видах водорослей защищенных бухт [Ремизова, 2008].
Весной в мейобентосе обнаружено 16 таксономических групп, среди которых к эумейобентосу относятся: Turbellaria, Nematoda, Harpacticoida, Ostracoda, Foraminifera, Aearina к псевдомейобентосу-Polychaeta, Oligochaeta, Cumacea, Anisopoda, Isopoda, Amphipoda, Loricata, MytilidaeBivalvia, Gastropoda, Chironomidae (Insecta) [Ремизова, 2008].
Мейобентос Черного моря, как объект изучения,
представляет особый интерес. Во-первых, это связано с тем, что фауна моря имеет
различное происхождение, отражающее геологическое прошлое водоема [Гусева,
1995]. В составе бентических форм (включая мейобентос) встречаются понтические
(каспийские) реликты, виды бореально-атлантического происхождения,
средиземноморские иммигранты и, наконец, экзотические виды, случайно завезенные
судами из других районов Мирового океана [Зайцев, Александров, Миничева, 2006].
Во-вторых, в силу ряда природных и антропогенных причин (высокая изоляция от
Мирового океана, обширная водосборная площадь, замедленное вертикальное
перемешивание водных масс, индустриализация прибрежной зоны, эвтрофирование и
т. д.) в Черном море произошли глубокие биотопические изменения, отразившиеся
на их обитателях. В последние годы этот аспект привлек к себе внимание не
только местных специалистов, но и международных организаций, включая структуры
ООН [Александров, 2004].
2. Физико-географическая и гидрохимическая
характеристика Новороссийской бухты
Для Черного моря, особенно Кавказского побережья, характерна небольшая изрезанность берегов.
Новороссийская бухта образовалась во время вторжения моря в сушу при разломе и опускании одного из прибрежных горных хребтов. Она представляет собой естественный постоянный водоём, заполненный морской водой [Тильба, Мнацеканов, Крутолапов, 2006]. Его западные берега не образуют поднятий, и протянувшаяся вдоль береговой полосы Суджукская коса сформировала небольшую лагуну с мелководными участками. Восточные берега бухты представляют собой высокие каменно-земляные обрывы с узкой пляжной полосой. Внутренняя часть бухты обрамлена набережными, к которым подступают городские кварталы. Угодье является крупным морским портом, снабжено выдающимися далеко в море молами, пристанями [Маккавеева, 1979].
Новороссийская бухта, известная также под названием Суджукской (по названию турецкой крепости Суджук-Кале) и Цемесской (по названию впадающей в нее реки Цемесс), является самой обширной и глубоководной на Кавказском побережье. Карта-схема Новороссийской бухты представлена на рисунке 1. Длина бухты 15 км, ширина у входа 9 км, в средней части 4,6 км. [Харитонова, 1981]. Береговая полоса почти не изрезана. Восточный и западный берега значительно отличаются друг от друга. Восточный берег ограничен сплошным хребтом гор (хребет Мархот), круто спускающимся к бухте отвесными утесами. Этот берег разделен мысами Шесхарис и Пинай, примерно, на три равные части. Западный берег представляет собой невысокую, полого спускающуюся к морю холмистую местность [Маккавеева, 1979].
Рельеф дна бухты ровный, с преобладающими глубинами 21-27 м. У берега глубина резко уменьшается.
Расположение грунтов типичное для Черного моря. Вдоль берегов идет полоса скал, расширяющаяся по мере продвижения выхода из бухты на глубине от 5 до 25 м. На глубинах 5-15 м. и 19-25 м. лежит слой крупного ракушечника или песка [Харитонова, 1981].
По гидрологическому режиму Новороссийская бухта делится на три района: порт, средняя часть бухты и район выхода из бухты [Харитонова, 1981].
Порт - отгороженная молами вершина бухты. Для него характерна мелководность (10-12 м.) и значительная удаленность от вод открытого моря, это является одной из причин сильной загрязненности портовой части [Макавеева, 1979].
Средняя часть располагается от моловых
сооружений до мыса Пенай. Максимальная глубина 25 м. Для той части бухты
характерно широкое сообщение вод с открытым морем. Загрязнение городскими
стоками здесь меньше, но все же значительное [Харитонова, 1981].
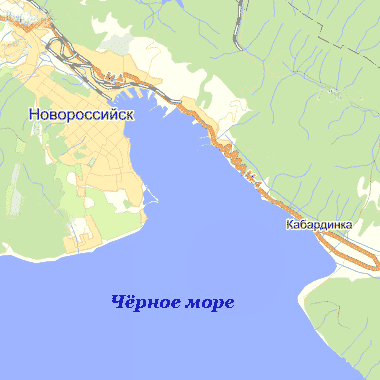
Рисунок 1 - Карта-схема Новороссийской бухты
Часть бухты, расположенная у выхода, имеет глубину до 30- 35 м. Гидрологический режим определяется гидрологическим режимом прибрежной полосы северо-восточной части Черного моря, отличный от условий открытого моря [Тильба, Мнацеканов, Крутолапов, 2006].
К концу ҲҲ столетия экологическое состояние Черного моря учеными и специалистами было определено как кризисное. Это не могло не вызвать обеспокоенность мирового сообщества и проживающих на его берегах народов.
К основным экологическим проблемам Черного моря на данный момент относят:
. эвтрофикация (цветение воды). Из-за попадания в прибрежные акватории остатков химических удобрений и плохо очищенных сточных вод;
. загрязнение воды нефтепродуктами и другими токсическими веществами;
. биологическое загрязнение чужеродными видами (рапана, гребневик);
. уменьшение биологического разнообразия, выраженное в снижении численности или полном исчезновении черноморских видов флоры и фауны;
. сокращение площади природных ландшафтов, лесных и особо охраняемых территорий в прибрежных зонах;
. накопление твердых бытовых отходов [Долинский, Патлатюк,2013].
Площадь поверхности Черного моря в шесть раз меньше площади его водосбора. По этой причине, состояние Черного моря сильно зависит от хозяйственной деятельности на суше, как прибрежных, так и других государств его бассейна. На качество прибрежных вод Черного моря влияют береговые источники антропогенного загрязнения и в первую очередь большие населенные пункты на побережье [Глумов, Кочеткова,1966]. Вследствие деятельности объектов коммунального хозяйства населенных пунктов в морские воды поступает значительная часть загрязняющих веществ. В последнее время в связи с усилением природоохранной деятельности, снижения объемов сбросов и улучшением качества очистки сточных вод в регионе наметилась четкая тенденция по снижению поступления загрязняющих веществ в море. К сожалению, мер, которые предпринимают черноморские государства недостаточно [Вершинин, 2003].
Одна из самых серьезных экологических проблем Черного моря заключается в том, что впадающие в него реки несут с собой не только пестициды или тяжелые металлы, но и азот и фосфор, в больших количествах смывающиеся с полей.
Основным международным документом, регулирующим
вопросы охраны Черного моря, является Конвенция о защите Черного моря от
загрязнения, подписанная шестью черноморскими странами - Болгарией, Грузией,
Россией, Румынией, Турцией и Украиной в 1992 году в Бухаресте [Долинский,
Патлатюк, 2004].
3. Материал и методы исследования
Материал для данной работы был собран в период с 23.06.2014 г. по 20.04.2014 г. в Новороссийской бухте на базе Новороссийского учебного и научно-исследовательского морского биологического центра Кубанского государственного университета.
Пробы мейобентоса отбирались в акватории Новороссийской бухты в трех различных точках: «Яхтклуб», «Рыбзавод», «поселок Алексино». При выборе станций для отбора проб основным критерием была непосредственная близость или удаленность от акватории порта. Станция «Яхтклуб» находится ближе всего к акватории порта, за ней идет станция «Рыбзавод», «поселок Алексино» находится в открытой части бухты.
Пробы отбирались два раза (01.07.2014 и 09.07.2014) в урезовой зоне (0,5м).
С каждой точки было взято по шесть проб. Станции отбора проб показаны на рисунке 2.
Сбор и обработку материала проводили по общепринятым методикам.
При сборе материала таллом водорослей мы накрывали мешком из мельничного газа № 49, подрезали подошву, затем затягивали отверстие мешка и выносили его на поверхность.
Каждый таллом водорослей нами этикетировался и помещался в полиэтиленовый мешок, и сутки пробы выдерживались без фиксации для более полного отделения животных от водорослей. Пробы промывали пресной проточной водой в большой емкости.
Смыв процеживали через систему сит (верхнее сито
- макрозообентосное, с ячеей 1 мм, нижнее, с ячеей 0,1 мм - мейозообентосное),
после чего пробы фиксировали 4% раствором формалина [Маккавеева, 1979].
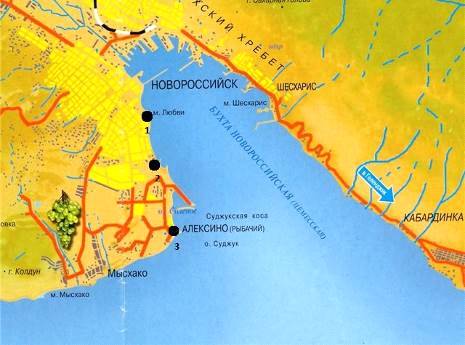
- Яхтклуб; 2 - Рыбзавод; 3 - поселок Алексино
Рисунок 2 - Станции отбора проб
Водоросли идентифицировались нами до вида, определялась их биомасса с целью последующего пересчета мейофауны на 1 кг массы водоросли. Для пересчета численности мейобентоса на 1 м2 дна мы использовали данные по общему проективному покрытию.
Разбор проб проводился в камере Богорова под бинокуляромМБС-10 при увеличении 8×2. Произвели подсчет основных групп мейобентоса [Маккавеева, 1979]. При расчете биомассы мейобентоса используются данные по средним весам, полученные для отдельных групп мейобентоса. Для расчета биомассы отдельных крупных мейобентосных организмов используются номограммы Численко [Численко, 1968].
Для идентификации водорослей был использован
определитель зеленых, бурых и красных водорослей южных морей СССР [Зинова,
1967]. Для идентификации организмов, обнаруженных в пробах, был использован
определитель фауны Черного и Азовского морей [Баранова, Бэческу, Голиков,
1986]. Также в работе было проведено сравнение сходства таксономического
состава на разных станциях отбора проб. Для этого использовался коэффициент
Серенсена, формула для расчета данного коэффициента выглядит таким образом
(формула 1):