Материал: Меледина Т.В., Давыденко С.Г. Дрожжи Saccharomyces cerevisiae. Морфология, химический состав, метаболизм
ко 1/4 митохондриальных белков. Основная часть белков митохондрий синтезируется на цитоплазматических рибосомах.
В митохондриях проходят важнейшие биохимические процессы, связанные с дыханием. Большая часть ферментов цикла трикарбоновых кислот (ЦТК) находится в матриксе митохондрии. Ферменты, участвующие в транспорте электронов и окислительном фосфорилировании, локализованы на внутренней мембране митохондрии, в том числе и на кристах.
На развитие митохондрий влияют не только физико-химические условия культивирования (рН, температура, концентрация кислорода
ит.д.), но и состав питательной среды. Так, добавление липидов, например олеиновой кислоты или эргостерола, приводит к образованию крист. На развитие митохондрии оказывает влияние также концентрация глюкозы в среде, которая вызывает проявление эффекта Кребтри. Активность митохондрий, связанная с ферментами ЦТК, зависит от наличия в среде культивирования факторов роста, макро- и микроэлементов и стимуляторов биосинтетических процессов. Так, при отсутствии в среде биотина снижается интенсивность процесса образования оксалоацетата, в результате чего замедляется работа цикла
иуменьшается потребность в дыхательных ферментах, следовательно, снижается потребность в развитых мембранных структурах, на которых они локализуются. Поэтому снижается скорость деления митохондрий и уменьшается содержание в них крист.
Эндоплазматическая сеть (ЭПC), или эндоплазматический ре-
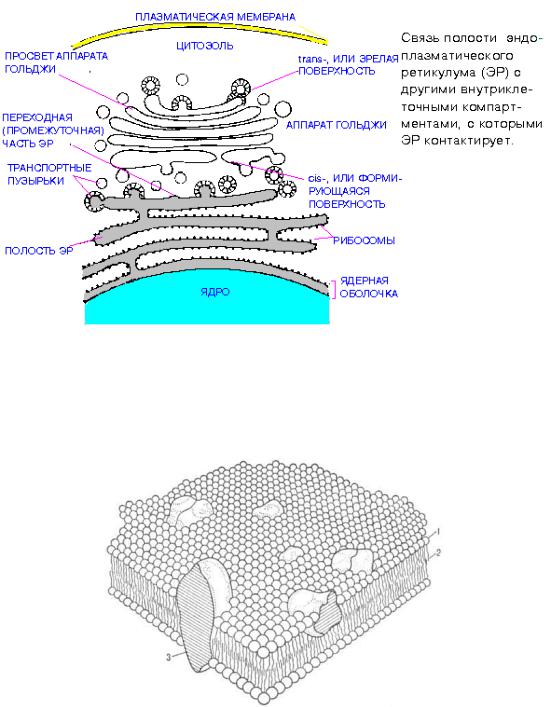
тикулум (ЭПР), – это система канальцев, цистерн и пузырьков, которые пронизывают всю протоплазму клетки и соединены друг с другом и другими органеллами, в частности с ядром и рибосомами (рис. 3.5). От степени разветвленности этой сети зависит активность многих метаболических процессов, в частности синтез белков, жиров
иуглеводов. ЭПР развита больше в аэробных дрожжах, чем в анаэробных, а также в молодых клетках больше, чем в старых.
ЭПР представляет собой трехслойную липопротеидную мембрану, на которой находятся многочисленные ферменты (рис. 3.6). ЭПР обеспечивает синтез и передвижение различных метаболитов в клетке и играет роль временных хранилищ выработанных продуктов. Например, белки, синтезированные в рибосомах, проходят в каналы ЭПР и по ним переносятся в те участки клетки, для которых они
21
предназначены. Во время перемещения белок может фосфорилироваться или превращаться в гликопротеид.
Рис. 3.5. Связь ЭПР с другими органеллами клеток
Таким образом, основными функциями ЭПР являются синтез белка, модификация белков, а также синтез липидов.
Рис. 3.6. Схема липопротеидной мембраны: 1, 2 – мембрана; 3 – белок
22
Эндоплазматический ретикулум у дрожжей делится на шероховатый и гладкий. В первом случае на поверхности мембраны, обращенной к цитоплазме, находятся рибосомы, на которых идет синтез белка. Гладкий ретикулум (ГР) является производным от шероховатого. Он участвует в синтезе липидов, необходимых для формирования всех клеточных мембран, а также углеводов. Кроме того, ГР связывает шероховатый ретикулум с аппаратом Гольджи.
Морфология ЭПР зависит от условий культивирования, фазы роста дрожжей и физиологического состояния клеток. Интенсивно растущие клетки имеют хорошо развитый ретикулум, поэтому его можно обнаружить только с помощью электронного микроскопа. Однако по мере снижения метаболической активности дрожжей происходит уменьшение поверхности мембран и их разветвленности, в результате появляются вакуоли. Таким образом, наблюдается взаимосвязь между структурой ЭПР и физиологическим состоянием дрожжей.
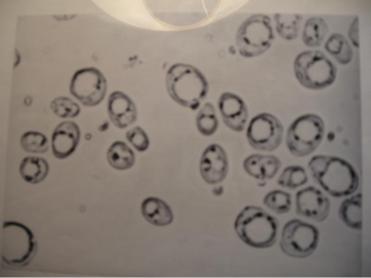
Процесс возникновения вакуолей представляется следующим образом: канальцы и микроцистерны ЭПР утолщаются, соединяются друг с другом, образуя более крупные полости (пузырчатая вакуоль), которые постепенно становятся все крупнее (рис. 3.7).
Рис. 3.7. Фотография дрожжей, содержащих вакуоли различного размера (увеличение 900 Х). Темные гранулы, локализованные на поверхности вакуолей, – это жировые включения
23
Представление о том, что вакуоли являются производными других мембранных систем клетки, подтверждается фактом их возрастного образования в клетках. При внесении дрожжей в свежую питательную среду эти вакуоли исчезают, а содержащиеся в них вещества используются в биосинтетических процессах. Ввиду активизации обмена веществ в клетках вновь возникает необходимость в хорошо развитой ЭПР сети, необходимой для синтеза и транспорта новых компонентов к различным органоидам клетки. Поэтому уже в конце лаг-фазы роста популяции эта сеть развита и имеет значительную протяженность. Контролирует синтез новых мембран ядро. В фазе замедления роста вновь происходят морфологические изменения ЭПР, и в стационарной фазе уже можно наблюдать появление вакуоли.
Роль вакуоли, прежде всего, заключается в поддержании внутриклеточного давления. Осмотическая регуляция в клетке происходит постоянно, только в молодом возрасте ее осуществляют эндоплазматическая сеть и аппарат Гольджи, а в зрелом – вакуоль.
В вакуоли скапливаются низкомолекулярные продукты гидролиза, например свободные аминокислоты, липиды, волютин, минеральные вещества
Аппарат Гольджи. Значительная часть гладких мембран отделена от ЭПР и организована в самостоятельные структурные образования, которые называются комплексом Гольджи, состоящим из системы пузырьков и дисков, соединенных друг с другом (см. рис. 3.5).
Основными функциями аппарата Гольджи являются модификация, накопление, сортировка и направление различных веществ в соответствующие внутриклеточные компартменты (отделения), а также за пределы клетки.
Аппарат Гольджи принимает участие в построении клеточной мембраны, в формировании лизосом, вакуолей и хитосом, он имеет непосредственное отношение к секреции белков и полисахаридов, а также выполняет функции транспорта секретируемых веществ в другие участки клетки и к ее поверхности.
Лизосомы – органеллы, окруженные однослойной мембраной, осуществляют «пищеварительные» функции. Они являются производными ЭПР и аппарата Гольджи. Различают первичные и вторичные лизосомы. В первичных лизосомах, локализованных в цитоплазме, ферменты находятся в неактивной форме. Их активность прояв-
24
ляется только тогда, когда они внедряются в вакуоль, образуя вторичную вакуоль.
Хитосомы грибов содержат фермент хитинсинтазу, которая катализирует синтез микрофибрилл хитина. Хитосомы переносят микрофибриллы к клеточной стенке, где происходит биосинтез рубцов.
Пероксисомы – органеллы, в которых аккумулированы окис- лительно-восстановительные ферменты, например, каталаза, которая восстанавливает пероксид водорода до воды. Эти структуры имеют функционально-морфологическую связь с митохондриями, так как ферменты, содержащиеся в митохондриях и пероксисомах, дополняют друг друга в метаболизме дрожжей. Прослежена связь этих органелл также с ЭПР, аппаратом Гольджи , ядром и др.
3.6. ЗАПАСНЫЕ ВЕЩЕСТВА
Помимо органелл в цитоплазме находятся запасные вещества – включения. Это волютин, липиды, гликоген, белки.
Волютин, или метахроматин. Название метахроматин связано с тем, что он вызывает характерное изменение цвета у красителя метиленового синего.
Волютин у дрожжей локализован в вакуолях. Это кислоторастворимое соединение, состоящее из полифосфата, белка, РНК, ионов Mg+2 и Ca+2. Волютин является резервом фосфора в клетке. Он накапливается при замедлении роста клеток, вызванного нехваткой питательных компонентов. Микроскопирование в обычном световом микроскопе не позволяет обнаружить волютин, так как он находится в вакуоли в аморфном состоянии, поэтому его определяют цитохимическим методом.
Помимо волютина в клетках содержатся высокополимерные полифосфаты (ПФ), локализованные на ЦПМ или вблизи от нее и обладающие большой метаболической активностью. Некоторое количество ПФ находится в клеточной оболочке. Именно эти ПФ принимают непосредственное участие в активном транспорте сахаров из окружающей среды в клетку и участвуют в синтезе маннана. Вакуолярная фракция ПФ метаболически малоактивна, и ее можно рас-
25