Материал: Курс занятий. Липиды
Переваривание нейтрального жира в желудочно-кишечном тракте. Липазы и их роль. Факторы, влияющие на степень гидролиза нейтрального жира.
У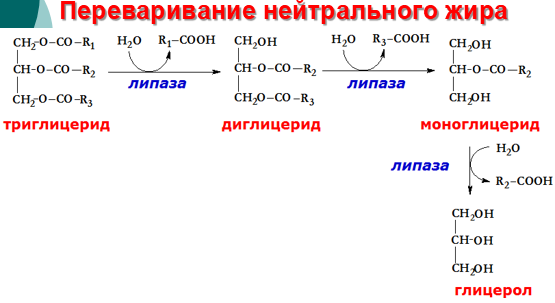 словия
переваривания липидов:
словия
переваривания липидов:
1. активность липазы (температура, оптимум рН, активаторы)
2. степень эмульгированности жира (наличие эмульгаторов)
 Жиры
составляют до 90% липидов, поступающих
с пищей. Переваривание жиров происходит
в тонком кишечнике, однако уже в желудке
небольшая часть жиров гидролизуется
под действием "липазы
языка".
Этот фермент синтезируется железами
на дорсальной поверхности языка и
относительно устойчив при кислых
значениях рН желудочного сока. Поэтому
он действует в течение 1-2 ч на жиры пищи
в желудке. Однако вклад этой липазы в
переваривание жиров у взрослых людей
незначителен. Основной процесс
переваривания происходит в тонкой
кишке.
Жиры
составляют до 90% липидов, поступающих
с пищей. Переваривание жиров происходит
в тонком кишечнике, однако уже в желудке
небольшая часть жиров гидролизуется
под действием "липазы
языка".
Этот фермент синтезируется железами
на дорсальной поверхности языка и
относительно устойчив при кислых
значениях рН желудочного сока. Поэтому
он действует в течение 1-2 ч на жиры пищи
в желудке. Однако вклад этой липазы в
переваривание жиров у взрослых людей
незначителен. Основной процесс
переваривания происходит в тонкой
кишке.
Так как жиры - нерастворимые в воде соединения, то они могут подвергаться действию ферментов, растворённых в воде только на границе раздела фаз вода/жир. Поэтому действию панкреатической липазы, гидролизующей жиры, предшествует эмульгирование жиров. Эмульгирование (смешивание жира с водой) происходит в тонком кишечнике под действием солей жёлчных кислот. Жёлчные кислоты синтезируются в печени из холестерола и секретируются в жёлчный пузырь. Содержимое жёлчного пузыря - жёлчь. Это вязкая жёлто-зелёная жидкость, содержащая главным образом жёлчные кислоты; в небольшом количестве имеются фосфолипиды и холестерол. Жёлчные кислоты представляют собой в основном конъюгированные жёлчные кислоты: таурохолевую, гликохолевую и другие. После приёма жирной пищи жёлчный пузырь сокращается и жёлчь изливается в просвет двенадцатиперстной кишки. Жёлчные кислоты действуют как детергенты, располагаясь на поверхности капель жира и снижая поверхностное натяжение. В результате крупные капли жира распадаются на множество мелких, т.е. происходит эмульгирование жира. Эмульгирование приводит к увеличению площади поверхности раздела фаз жир/вода, что ускоряет гидролиз жира панкреатической липазой. Эмульгированию способствует и перистальтика кишечника. Переваривание жиров - гидролиз жиров панкреатической липазой. Оптимальное значение рН для панкреатической липазы ≈8 достигается путём нейтрализации кислого содержимого, поступающего из желудка, бикарбонатом, выделяющимся в составе сока поджелудочной железы:
Н+ + НСО3- → Н2СО3 → Н2О + СО2 ↑.
Выделяющийся углекислый газ способствует дополнительному перемешиванию содержимого тонкой кишки.
Панкреатическая липаза выделяется в полость тонкой кишки из поджелудочной железы вместе с белком колипазой. Колипаза попадает в полость кишечника в неактивном виде и частичным протеолизом под действием трипсина превращается в активную форму. Колипаза своим гидрофобным доменом связывается с поверхностью мицеллы эмульгированного жира. Другая часть молекулы способствует формированию такой конформации панкреатической липазы, при которой активный центр фермента максимально приближен к своим субстратам - молекулам жиров поэтому скорость реакции гидролиза жира резко возрастает
Панкреатическая липаза гидролизует жиры преимущественно в положениях 1 и 3 поэтому основными продуктами гидролиза являются свободные жирные кислоты и 2-моноацилглицеролы (β-моноацилглицеролы). Молекулы 2-моноацилглицеролов также обладают детергентными свойствами и способствуют эмульгированию жира.
Переваривание фосфолипидов и стероидов в желудочно-кишечном тракте. Ферменты, роль.
У взрослого человека процесс переваривания и всасывания происходит в 12-перстной кишке при оптимальном рН=8 для действия ферментов.
У грудных детей основной пищей служит молоко, содержащее жиры в эмульгированном виде и переваривание липидов происходит в желудке, рН желудочного сока 5-5,5.
Жиры, проходя ЖКТ расщепляются до своих конечных продуктов.
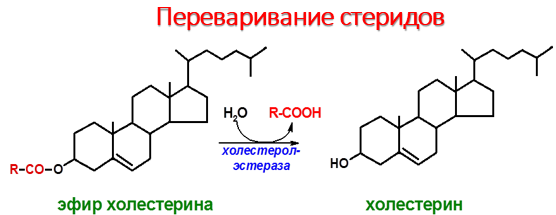
В процессе переваривания и всасывания принимают участие соли желчных кислот, синтезирующиеся в печени из ХС и при этом конъюгируются с таурином или глицином, образуя парные желчные кислоты, например, таурохолевую, гликохолевую.Подавляющая часть фосфолипидов содержимого тонкой кишки приходится на фосфатидилхолин (лецитин), основная масса которого поступает в кишечник с желчью (11–12 г/сут) и меньшая часть (1–2 г/сут) – с пищей. Существует две точки зрения относительно судьбы поступивших в тонкую кишку экзогенных и эндогенных фосфолипидов. Согласно одной из них, и те, и другие фосфолипиды подвергаются в кишечнике атаке со стороны фосфолипазы А2, катализирующей гидролиз сложноэфирной связи в β-положении. В результате катализируемой фосфолипазой А2 реакции глицерофосфолипиды расщепляются с образованием лизофосфолипида и жирной кислоты.
Л изофосфолипид
может подвергаться расщеплению при
действии другого фермента панкреатического
сока – лизофосфолипазы. В результате
из лизолецитина освобождается последняя
частица жирной кислоты и образуется
глицерофосфохолин, который хорошо
растворяется в водной среде и всасывается
из кишечника в кровь. Сторонники другой
точки зрения считают, что фосфолипиды
«желчного» (более точно печеночного)
происхождения в отличие от пищевых
фосфолипидов не подвергаются воздействию
фосфолипазы А2. Следовательно, функция
«желчных» фосфолипидов исключительно
связана с гепатоэнтеральной циркуляцией
желчи: с желчью они поступают в кишечник,
с желчными кислотами участвуют в
мицеллярной солюбилизации липидов и
вместе с ними возвращаются в печень.
Таким образом, существует как бы два
пула фосфолипидов в кишечнике: «желчный»,
защищенный от действия фосфолипазы А2,
и «пищевой», подверженный ее действию.
изофосфолипид
может подвергаться расщеплению при
действии другого фермента панкреатического
сока – лизофосфолипазы. В результате
из лизолецитина освобождается последняя
частица жирной кислоты и образуется
глицерофосфохолин, который хорошо
растворяется в водной среде и всасывается
из кишечника в кровь. Сторонники другой
точки зрения считают, что фосфолипиды
«желчного» (более точно печеночного)
происхождения в отличие от пищевых
фосфолипидов не подвергаются воздействию
фосфолипазы А2. Следовательно, функция
«желчных» фосфолипидов исключительно
связана с гепатоэнтеральной циркуляцией
желчи: с желчью они поступают в кишечник,
с желчными кислотами участвуют в
мицеллярной солюбилизации липидов и
вместе с ними возвращаются в печень.
Таким образом, существует как бы два
пула фосфолипидов в кишечнике: «желчный»,
защищенный от действия фосфолипазы А2,
и «пищевой», подверженный ее действию.
Всасывание продуктов переваривания липидов.
Продукты гидролиза липидов - жирные кислоты с длинным углеводородным радикалом, 2-моноацилглицеролы, холестерол, а также соли жёлчных кислот образуют в просвете кишечника структуры, называемые смешанными мицеллами. Смешанные мицеллы построены таким образом, что гидрофобные части молекул обращены внутрь мицеллы, а гидрофильные - наружу, поэтому мицеллы хорошо растворяются в водной фазе содержимого тонкой кишки. Стабильность мицелл обеспечивается в основном солями жёлчных кислот. Мицеллы сближаются со щёточной каймой клеток слизистой оболочки тонкого кишечника, и липидные компоненты мицелл диффундируют через мембраны внутрь клеток. Вместе с продуктами гидролиза липидов всасываются жирорастворимые витамины A, D, Е, К и соли жёлчных кислот. Наиболее активно соли жёлчных кислот всасываются в подвздошной кишке. Жёлчные кислоты далее попадают через воротную вену в печень, из печени вновь секретируются в жёлчный пузырь и далее опять участвуют в эмульгировании жиров. Этот путь жёлчных кислот называют "энтерогепатическая циркуляция". Каждая молекула жёлчных кислот за сутки проходит 5- 8 циклов, и около 5% жёлчных кислот выделяется с фекалиями.
Всасывание жирных кислот со средней длиной цепи, образующихся, например, при переваривании липидов молока, происходит без участия смешанных мицелл. Эти жирные кислоты из клеток слизистой оболочки тонкого кишечника попадают в кровь, связываются с белком альбумином и транспортируются в печень.
О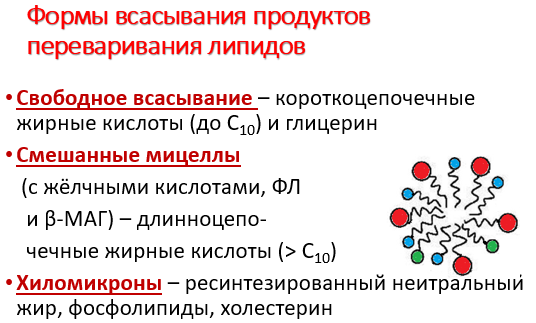 бразование
смешанных мицелл и всасывание продуктов
гидролиза.
Продукты
гидролиза липидов - жирные кислоты с
длинным углеводородным радикалом,
2-моноацилглицеролы, холестерол, а также
соли жёлчных кислот образуют в просвете
кишечника структуры, называемые
смешанными мицеллами. Смешанные мицеллы
построены таким образом, что гидрофобные
части молекул обращены внутрь мицеллы,
а гидрофильные - наружу, поэтому мицеллы
хорошо растворяются в водной фазе
содержимого тонкой кишки. Стабильность
мицелл обеспечивается в основном солями
жёлчных кислот. Мицеллы сближаются со
щёточной каймой клеток слизистой
оболочки тонкого кишечника, и липидные
компоненты мицелл диффундируют через
мембраны внутрь клеток.
бразование
смешанных мицелл и всасывание продуктов
гидролиза.
Продукты
гидролиза липидов - жирные кислоты с
длинным углеводородным радикалом,
2-моноацилглицеролы, холестерол, а также
соли жёлчных кислот образуют в просвете
кишечника структуры, называемые
смешанными мицеллами. Смешанные мицеллы
построены таким образом, что гидрофобные
части молекул обращены внутрь мицеллы,
а гидрофильные - наружу, поэтому мицеллы
хорошо растворяются в водной фазе
содержимого тонкой кишки. Стабильность
мицелл обеспечивается в основном солями
жёлчных кислот. Мицеллы сближаются со
щёточной каймой клеток слизистой
оболочки тонкого кишечника, и липидные
компоненты мицелл диффундируют через
мембраны внутрь клеток.
Желчные кислоты, строение, роль в переваривании и всасывании липидов.
П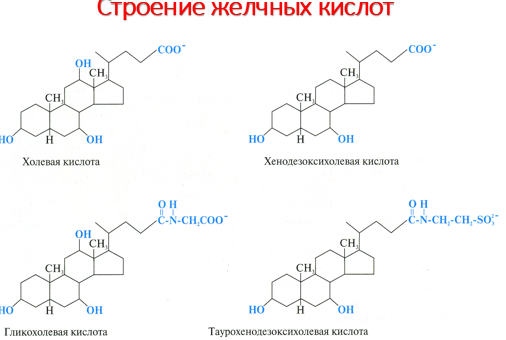 осле
того как химус попадает в двенадцатиперстную
кишку, здесь прежде всего происходит
нейтрализация соляной кислоты желудочного
сока, попавшей в кишечник с пищей,
бикарбонатами, содержащимися в
панкреатическом и кишечном соках.
Выделяющиеся при разложении бикарбонатов
пузырьки углекислого газа способствуют
хорошему перемешиванию пищевой кашицы
с пищеварительными соками. Одновременно
начинается эмульгирование жира. Наиболее
мощное эмульгирующее действие на жиры,
несомненно, оказывают соли желчных
кислот, попадающие в двенадцатиперстную
кишку с желчью в виде натриевых солей,
большая часть которых конъюгирована с
глицином или таурином. Желчные кислоты
представляют собой основной конечный
продукт обмена холестерина.
осле
того как химус попадает в двенадцатиперстную
кишку, здесь прежде всего происходит
нейтрализация соляной кислоты желудочного
сока, попавшей в кишечник с пищей,
бикарбонатами, содержащимися в
панкреатическом и кишечном соках.
Выделяющиеся при разложении бикарбонатов
пузырьки углекислого газа способствуют
хорошему перемешиванию пищевой кашицы
с пищеварительными соками. Одновременно
начинается эмульгирование жира. Наиболее
мощное эмульгирующее действие на жиры,
несомненно, оказывают соли желчных
кислот, попадающие в двенадцатиперстную
кишку с желчью в виде натриевых солей,
большая часть которых конъюгирована с
глицином или таурином. Желчные кислоты
представляют собой основной конечный
продукт обмена холестерина.
Роль желчных кислот
Эмульгируют жиры
Активируют панкреатическую липазу.
Участвуют во всасывании продуктов распада липидов.
После всасывания продуктов гидролиза жиров в стенке кишечника происходит ресинтез жиров: ТАГ, ЭХС, ФЛ.
Ресинтез триацилглицеринов в энтероцитах. Ресинтез фосфолипидов.
Р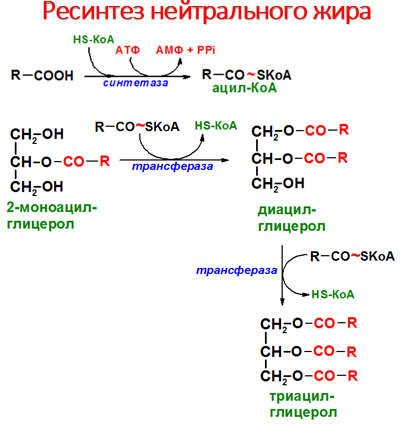 есинтез
жиров в слизистой оболочке тонкого
кишечника.
После
всасывания продуктов гидролиза жиров
жирные кислоты и 2-моноацилглицеролы в
клетках слизистой оболочки тонкого
кишечника включаются в процесс ресинтеза
с образованием триацилглицеролов Жирные
кислоты вступают в реакцию этерификации
только в активной форме в виде производных
коэнзима А, поэтому первая стадия
ресинтеза жиров - реакция активации
жирной кислоты:
есинтез
жиров в слизистой оболочке тонкого
кишечника.
После
всасывания продуктов гидролиза жиров
жирные кислоты и 2-моноацилглицеролы в
клетках слизистой оболочки тонкого
кишечника включаются в процесс ресинтеза
с образованием триацилглицеролов Жирные
кислоты вступают в реакцию этерификации
только в активной форме в виде производных
коэнзима А, поэтому первая стадия
ресинтеза жиров - реакция активации
жирной кислоты:
Р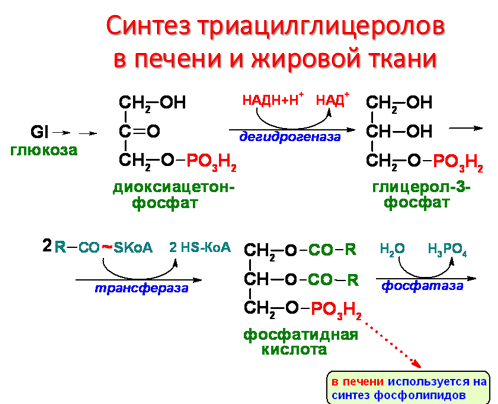 еакция
катализируется ферментом ацил-КоА-синтетазой
(тиокиназой). Затем ацил~КоА участвует
в реакции этерификации 2-моноацилглицерола
с образованием сначала диацилгли-церола,
а затем триацилглицерола. Реакции
ресинтеза жиров катализируют
ацилтранеферазы. В реакциях ресинтеза
жиров участвуют, как правило, только
жирные кислоты с длинной углеводородной
цепью. В ресинтезе жиров участвуют не
только жирные кислоты, всосавшиеся из
кишечника, но и жирные кислоты,
синтезированные в организме, поэтому
по составу ре-синтезированные жиры
отличаются от жиров, полученных с пищей.
Однако возможности "адаптировать"
в процессе ресинтеза состав пищевых
жиров к составу жиров организма человека
ограничены, поэтому при поступлении с
пищей жиров с необычными жирными
кислотами, например,
бараньего жира, в адипоцитах появляются
жиры, содержащие кислоты, характерные
для бараньего жира (насыщенные
разветвлённые жирные кислоты). В клетках
слизистой оболочки кишечника происходит
активный синтез глицерофосфолипидов,
необходимых для формирования структуры
липопротеинов - транспортных форм
липидов в крови.
еакция
катализируется ферментом ацил-КоА-синтетазой
(тиокиназой). Затем ацил~КоА участвует
в реакции этерификации 2-моноацилглицерола
с образованием сначала диацилгли-церола,
а затем триацилглицерола. Реакции
ресинтеза жиров катализируют
ацилтранеферазы. В реакциях ресинтеза
жиров участвуют, как правило, только
жирные кислоты с длинной углеводородной
цепью. В ресинтезе жиров участвуют не
только жирные кислоты, всосавшиеся из
кишечника, но и жирные кислоты,
синтезированные в организме, поэтому
по составу ре-синтезированные жиры
отличаются от жиров, полученных с пищей.
Однако возможности "адаптировать"
в процессе ресинтеза состав пищевых
жиров к составу жиров организма человека
ограничены, поэтому при поступлении с
пищей жиров с необычными жирными
кислотами, например,
бараньего жира, в адипоцитах появляются
жиры, содержащие кислоты, характерные
для бараньего жира (насыщенные
разветвлённые жирные кислоты). В клетках
слизистой оболочки кишечника происходит
активный синтез глицерофосфолипидов,
необходимых для формирования структуры
липопротеинов - транспортных форм
липидов в крови.
Ресинтез фосфолипидов в кишечной стенке. В энтероцитах наряду с ресинтезом триглицеридов происходит также и ресинтез фосфолипидов. В образовании фосфатидилхолинов и фосфатидилэтаноламинов участвует ресинтезированный диглицерид, а в образовании фосфатидилинозитолов – ресинтезированная фосфатидная кислота.
Образование хиломикронов и транспорт жиров.
Р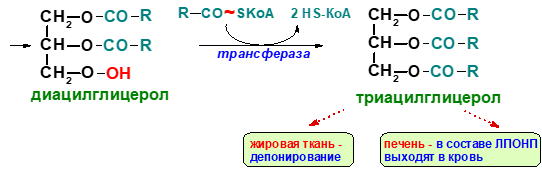
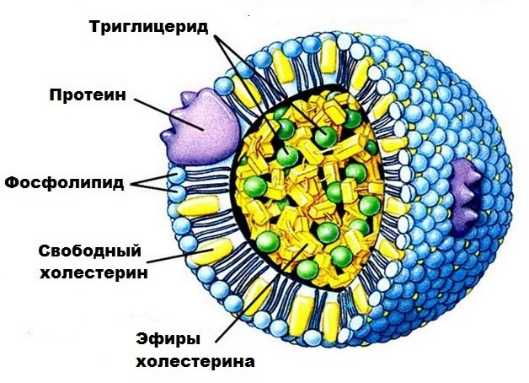 есинтезированные
в эпителиальных клетках кишечника
триглицериды и фосфолипиды, а также
поступивший в эти клетки из полости
кишечника холестерин (здесь он может
частично эстерифицироваться) соединяются
с небольшим количеством белка и образуют
относительно стабильные комплексные
частицы – хиломикроны (ХМ). ХМ- около 2%
белка, 7% фосфолипидов, 8% холестерина и
его эфиров и более 80% триглицеридов.
Диаметр ХМ от 0,1 до 5 мкм. Благодаря
большим размерам частиц ХМ не способны
проникать из эндотелиальных клеток
кишечника в капилляры и диффундируют
в лимфатическую систему кишечника, а
из нее – в грудной лимфатический проток.
Затем попадают в кровяное русло, т.е. с
их помощью осуществляется транспорт
экзогенных триглицеридов, холестерина
и частично фосфолипидов из кишечника
через лимфатическую систему в кровь
есинтезированные
в эпителиальных клетках кишечника
триглицериды и фосфолипиды, а также
поступивший в эти клетки из полости
кишечника холестерин (здесь он может
частично эстерифицироваться) соединяются
с небольшим количеством белка и образуют
относительно стабильные комплексные
частицы – хиломикроны (ХМ). ХМ- около 2%
белка, 7% фосфолипидов, 8% холестерина и
его эфиров и более 80% триглицеридов.
Диаметр ХМ от 0,1 до 5 мкм. Благодаря
большим размерам частиц ХМ не способны
проникать из эндотелиальных клеток
кишечника в капилляры и диффундируют
в лимфатическую систему кишечника, а
из нее – в грудной лимфатический проток.
Затем попадают в кровяное русло, т.е. с
их помощью осуществляется транспорт
экзогенных триглицеридов, холестерина
и частично фосфолипидов из кишечника
через лимфатическую систему в кровь
Вместе с продуктами гидролиза липидов всасываются жирорастворимые витамины A, D, Е, К и соли жёлчных кислот. Наиболее активно соли жёлчных кислот всасываются в подвздошной кишке. Жёлчные кислоты далее попадают через воротную вену в печень и вновь секретируются в жёлчный пузырь и далее опять участвуют в эмульгировании жиров. Этот путь жёлчных кислот называют "энтерогепатическая циркуляция".. Всасывание жирных кислот со средней длиной цепи, образующихся, например, при переваривании липидов молока, происходит без участия смешанных мицелл. Эти жирные кислоты из клеток слизистой оболочки тонкого кишечника попадают в кровь, связываются с белком альбумином и транспортируются в печень.
Нарушения переваривания и всасывания жиров. Нарушение переваривания жиров может быть следствием нескольких причин. Одна из них - нарушение секреции жёлчи из жёлчного пузыря при механическом препятствии оттоку жёлчи. Это состояние может быть результатом сужения просвета жёлчного протока камнями, образующимися в жёлчном пузыре, или сдавлением жёлчного протока опухолью, развивающейся в окружающих тканях. Уменьшение секреции жёлчи приводит к нарушению эмульгирования пищевых жиров и, следовательно, к снижению способности панкреатической липазы гидролизовать жиры.
Нарушение секреции сока поджелудочной железы и, следовательно, недостаточная секреция панкреатической липазы также приводят к снижению скорости гидролиза жиров. В обоих случаях нарушение переваривания и всасывания жиров приводит к увеличению количества жиров в фекалиях - возникает стеаторея (жирный стул). В норме содержание жиров в фекалиях составляет не более 5%. При стеаторее нарушается всасывание жирорастворимых витаминов (A, D, E, К) и незаменимых жирных кислот, поэтому при длительно текущей стеаторее развивается недостаточность этих незаменимых факторов питания с соответствующими клиническими симптомами. При нарушении переваривания жиров плохо перевариваются и вещества нелипидной природы, так как жир обволакивает частицы пищи и препятствует действию на них ферментов.
Л ипопротеинлипаза, ее роль.
Липопротеинлипаза — фермент, относящийся к классу липаз. ЛПЛ расщепляет триглицериды самых крупных по размеру и богатых липидами липопротеинов плазмы крови — хиломикронов и липопротеинов очень низкой плотности (ЛПОНП или ЛПНП)). ЛПЛ регулирует уровень липидов в крови, что определяет её важное значение в атеросклерозе.
Обмен нейтрального жира. Резервирование и мобилизация жиров в жировой ткани.
Жировой обмен — совокупность процессов переваривания и всасывания нейтральных жиров (триглицеридов) и продуктов их распада в желудочно-кишечном тракте, промежуточного обмена жиров и жирных кислот и выведение жиров, а также продуктов их обмена из организма. Понятия «жировой обмен» и «липидный обмен» часто используются как синонимы, т.к. входящие в состав тканей животных и растений входят нейтральные жиры и жироподобные соединения, объединяются под общим названием липиды. Жиры, как и гликоген, являются формами депонирования энергетического материала. Причем жиры - наиболее долговременные и более эффективные источники энергии. При голодании запасы жира у человека истощаются за 5-7 недель, тогда как гликоген полностью расходуется примерно за сутки. Если поступление жира превышает потребности организма в энергии, то жир депонируется в специализированных клетках жировой ткани. Кроме того, если количество поступающих углеводов больше, чем надо для депонирования в виде гликогена, то часть глюкозы также превращается в жиры. Таким образом, жиры в жировой ткани накапливаются в результате трех процессов:
1. поступают из экзогенных жиров пищи
2. поступают из эндогенных жиров, синтезированных в печени из глюкозы
3. образуются из глюкозы в самих клетках жировой ткани.
В организм взрослого человека с пищей ежесуточно поступает в среднем 70 г жиров животного и растительного происхождения. Здесь нет смысла раскрывать сложные процессы преобразований поступивших с пищей жиров с участием различных ферментов и желчной кислоты. Достаточно сказать, что конечным итогом этих преобразований является образование триглицеридов жировой ткани. В первом и втором случае этот синтез может протекать только в присутствии глюкозы. Инсулин стимулирует этот синтез, потому что в его присутствии повышается проницаемость мембран клеток жировой ткани для глюкозы. Т.е. процесс усвоения экзогенных и эндогенных жиров в жировой ткани является инсулинозависимым процессом. Проще говоря, для того, чтобы съеденный шмат сала правильно «улегся», нужна еще и глюкоза и нужен инсулин, чтобы она попала в клетки жировой ткани. При этом не имеет значения, откуда будет взята эта глюкоза. Из куска хлеба, съеденного с салом или из внутренних депо организма (при употреблении сала без хлеба). Т.е. процесс образования триглицеридов происходит с участием глюкозы. Той самой, уровень содержания которой регулируется автоматикой с участием того же вездесущего инсулина. Т.е. процесс усвоения и трансформации пищевых жиров происходит в присутствии инсулина и является инсулинозависимым процессом. Ну а третий процесс (процесс образования жиров из глюкозы) сам по себе является инсулинзависимым, поскольку для проникновения глюкозы в клетки инсулинзависимой жировой ткани инсулин нужен по определению.
В жировой ткани человека содержится большое количество жира, преимущественно в виде триглицеридов, которые выполняют в обмене жиров такую же функцию, как гликоген печени в обмене углеводов. Запасы триглицеридов могут потребляться при голодании, физической работе и других состояниях, требующих большой затраты энергии.
Мобилизацию депонированных жиров (липолиз) катализирует фермент под названием липаза. В результате жиры распадаются на глицерин и свободные жирные кислоты, которые из жировой ткани поступают в кровоток. Судьба жирных кислот в самой жировой ткани во многом зависит от содержания в ней глюкозы или, что более точно, от интенсивности протекающего в ней гликолиза. При голодании, когда содержание глюкозы в жировой ткани понижено, освободившиеся жирные кислоты не могут быть использованы жировой тканью для ресинтеза триглицеридов, и поэтому они быстро покидают эту ткань. Т.о., активация гликолиза в жировой ткани является фактором, способствующим накоплению в ней триглицеридов, а угнетение гликолиза, наоборот, способствует их удалению. Глицерин транспортируется в печень, где используется в реакциях глюконеогенеза. Жирные кислоты транспортируются кровью в разные органы и ткани, где включаются в процесс окисления. Другими словами, жировой обмен тесно связан с углеводным обменом и инсулинозависим. Дефицит инсулина приводит к ослаблению накопления жира в жировых депо и активизирует их расщепление на глицерин и жирные кислоты.