Материал: Курс занятий. Белки
Понятие о свободном амином азоте. Принцип метода определения, диагностическое значение.
Свободный аминный азот сыворотки крови - азот свободных аминокислот, содержащихся в сыворотке крови (в основном глу и асп)
Принцип метода количественного определения основан на способности нингидрина взаимодействовать с продуктами деградации аминокислот и давать сложный комплекс сине-фиолетового цвета. Интенсивность развивающегося окрашивания пропорциональна количеству свободных аминокислот. Определение проводятся в 3 этапа:
1. Осаждение белков, при этом сыворотка освобождается от α-аминного азота белковых молекул.
2. Реакция свободных аминокислот с нингидрином.
3. Электрофотоколориметрия и расчет.
Увеличение содержания свободных аминокислот в крови имеет ме-сто при деструктивных состояниях: печеночной коме, острой желтой атрофии печени, при отравлениях (фосфором, фенилгидразином, СС14, хлороформом), при квашиоркоре, тяжелых ожогах, шоке, истощающих поносах, после кровотечений. Уменьшение уровня аминокислот характерно для почечных поражений, для гиперпродукции инсулина, гормона роста, андрогенов
Трансметилирование. Метионин и s-аденозилметионин. Роль метионина в биосинтезе биологически важных соединений, в реакциях обезвреживания.
Трансметилирование (син. переметилирование)– реакции переноса метильной группы от донора к акцептору
М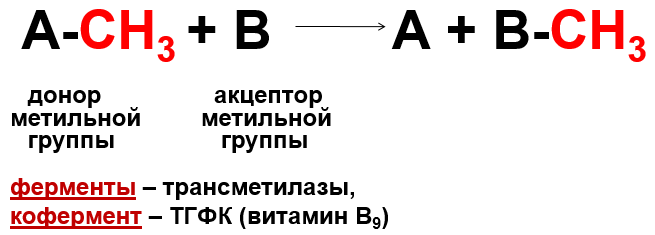 етионин -
незаменимая аминокислота, участвует в
реакциях дезаминирования, является
источником атома серы для синтеза
цистеина. Метионил-тРНК участвует в
инициации процесса трансляции. Метальная
группа метионина - мобильный одноуглеродный
фрагмент, используемый для синтеза ряда
соединений. Перенос метильной группы
метионина на соответствующий акцептор
называют реакцией трансметилирования.
Метальная группа в молекуле метионина
прочно связана с атомом серы, поэтому
непосредственным донором этого
одноутлеродного фрагмента служит
активная форма аминокислоты.
етионин -
незаменимая аминокислота, участвует в
реакциях дезаминирования, является
источником атома серы для синтеза
цистеина. Метионил-тРНК участвует в
инициации процесса трансляции. Метальная
группа метионина - мобильный одноуглеродный
фрагмент, используемый для синтеза ряда
соединений. Перенос метильной группы
метионина на соответствующий акцептор
называют реакцией трансметилирования.
Метальная группа в молекуле метионина
прочно связана с атомом серы, поэтому
непосредственным донором этого
одноутлеродного фрагмента служит
активная форма аминокислоты.
Реакция активация метионина Активной формой метионина является S-аденозилметионин (SAM) - сульфониевая форма аминокислоты, образующаяся в результате присоединения метионина к молекуле аденозина. Аденозин образуется при гидролизе АТФ. Эту реакцию катализирует фермент метионин аденозилтрансфераза, присутствующий во всех типах клеток. Структура (-S+-CH3) в SAM - нестабильная группировка, определяющая высокую активность метильной группы (отсюда термин "активный метионин"). Эта реакция уникальна для биологических систем, так как, по-видимому, является единственной известной реакцией, в результате которой освобождаются все три фосфатных остатка АТФ. Отщепление метильной группы от SAM и перенос её на соединение-акцептор катализируют ферменты метилтрансферазы. SAM в ходе реакции превращается в S-аденозилгомоцистеин (SAT)
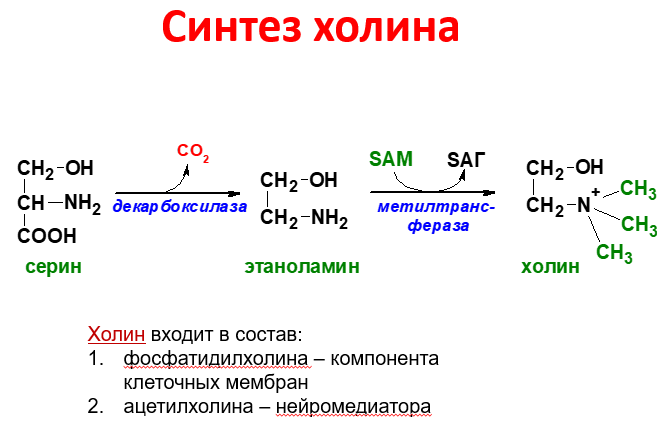
Синтез фосфатидилхолина из фосфатидилэтиноламина
Ф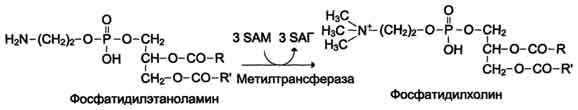 осфатидилхолины
(лецитины) - наиболее распространённая
группа глицерофосфолипидов, участвующих
в образовании мембран клеток и
липопротеинов, в составе которых
осуществляется транспорт липидов
осфатидилхолины
(лецитины) - наиболее распространённая
группа глицерофосфолипидов, участвующих
в образовании мембран клеток и
липопротеинов, в составе которых
осуществляется транспорт липидов
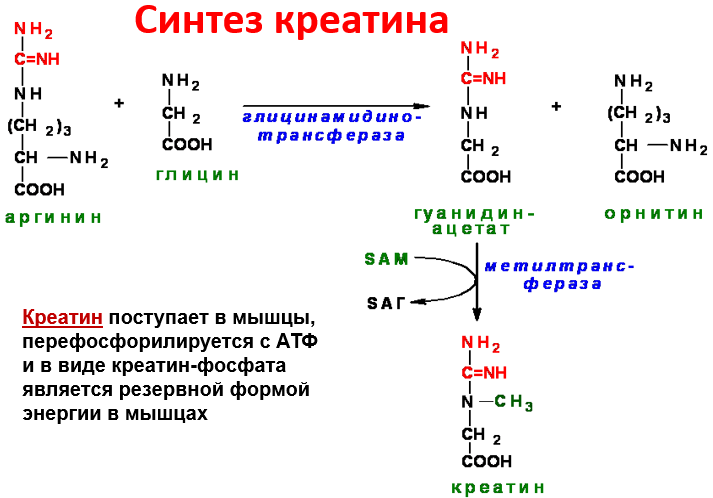
С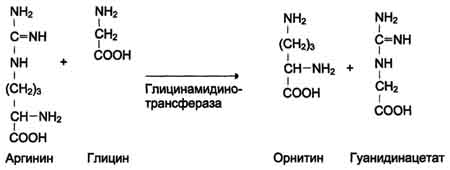 интез
креатина.Креатин
необходим для образования в мышцах
высокоэнергетического соединения -
кре-атинфосфата. Синтез креатина идёт
в 2 стадии с участием 3 аминокислот:
аргинина, глицина и метионина. В
почках образуется
гуанидинацетат при действии
глицинамидинотрансферазы
интез
креатина.Креатин
необходим для образования в мышцах
высокоэнергетического соединения -
кре-атинфосфата. Синтез креатина идёт
в 2 стадии с участием 3 аминокислот:
аргинина, глицина и метионина. В
почках образуется
гуанидинацетат при действии
глицинамидинотрансферазы
З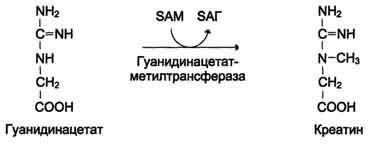 атем
гуанидинацетат транспортируется в
печень, где
происходит реакция его метилирования
атем
гуанидинацетат транспортируется в
печень, где
происходит реакция его метилирования
ГАМК в виде препаратов гаммалон или аминалон применяют при сосудистых заболеваниях головного мозга (атеросклероз, гипертония), нарушениях мозгового кровообращения, умственной отсталости, эндогенных депрессиях, травмах головного мозга, эпилепсии.
Д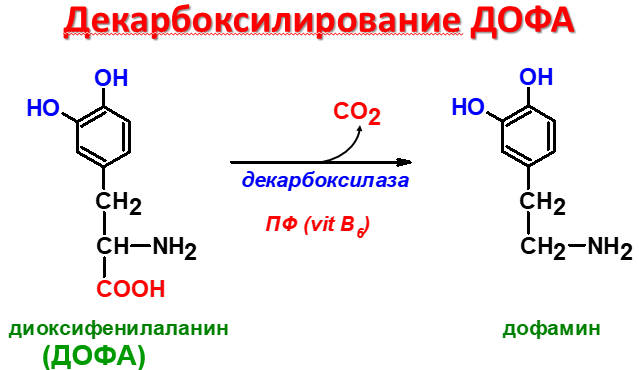
 офамин
– нейромедиатор, предшественник
норадреналина и адреналина
офамин
– нейромедиатор, предшественник
норадреналина и адреналина
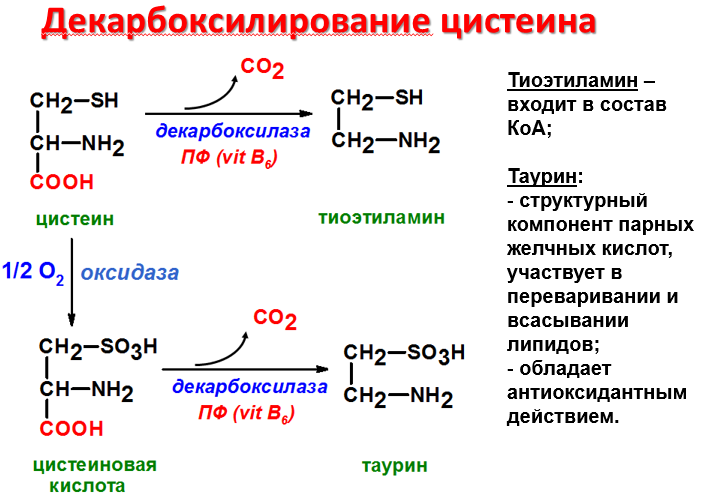
Функции аминов:
Амины, образовавшиеся при декарбоксилировании аминокислот, часто являются биологически активными веществами. Они выполняют функцию нейромедиаторов (серотонин, дофамин, ГАМК и др.), гормонов (норадреналин, адреналин), регуляторных факторов местного действия (гистамин, карнозин, спермин
Креатин с кровотоком переносится в мышцы и клетки мозга, где из него образуется высокоэнергетическое соединение - креатинфосфат. Определение содержания креатина и креати-нина в крови и моче используется для характеристики интенсивности работы мышц в спортивной медицине и при некоторых патологических состояниях. Определение активности фермента креатинкиназы и его изоферментных форм в крови используется в медицине для диагностики таких заболеваний, как инфаркт миокарда, миопатии, мышечные дистрофии и др.
Р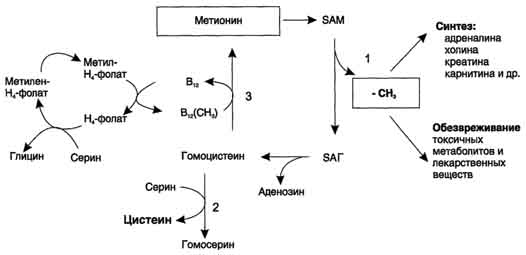 еакции
трансметилирования используются также
для:
еакции
трансметилирования используются также
для:
синтеза адреналина из норадреналина;
синтеза анзерина из карнозина;
метилирования азотистых оснований в нук-леотидах и др.;
инактивации метаболитов (гормонов, медиаторов и др.) и обезвреживания чужеродных соединений, включая и лекарственные препараты.
В организме метилированию могут подвергаться амины, фенолы и тиолы. В результате метилирования образуются соответствующие N-, О- и S-метильные конъюгаты. При метилировании чужеродных соединений и некоторых метаболитов переносчиком метильных групп является кофермент S-аденозилметионин. С участием метильных групп этого кофермента происходит метилирование перечисленных выше соединений. Реакции метилирования происходят под влиянием ферментных систем (метилтрансфераз)
Тетрагидрофолиевая кислота и ее роль в синтезе и переносе одноуглеродных радикалов.
В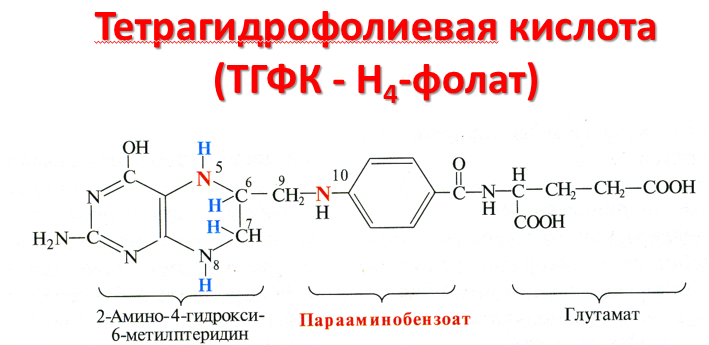 превращениях серина и глицина главную
роль играют ферменты, коферментами
которых служат производные фолиевой
кислоты. Этот витамин широко распространён
в животных и растительных пищевых
продуктах. Молекула фолиевой кислоты
(фолата) состоит из 3 частей: птеринового
производного, парааминобензойной и
глутаминовой кислот.
превращениях серина и глицина главную
роль играют ферменты, коферментами
которых служат производные фолиевой
кислоты. Этот витамин широко распространён
в животных и растительных пищевых
продуктах. Молекула фолиевой кислоты
(фолата) состоит из 3 частей: птеринового
производного, парааминобензойной и
глутаминовой кислот.
Г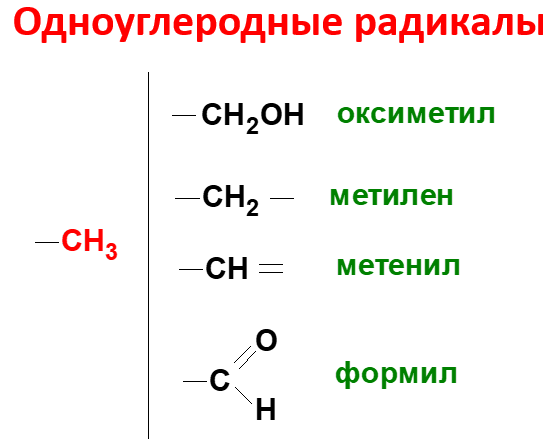 омоцистеин
может снова превращаться в метионин
под действием гомоцистеинметил-транс-феразы.
Донором метильной группы в этом случае
служит N5-метил-Н4-фолат:
омоцистеин
может снова превращаться в метионин
под действием гомоцистеинметил-транс-феразы.
Донором метильной группы в этом случае
служит N5-метил-Н4-фолат:
Образование и использование одноуглеродных фрагментов.Особое значение реакций катаболизма серина и глицина заключается в том, что они сопровождаются образованием одноуглеродного метиленового фрагмента (-СН2-). Метиленовая группа в молекуле метилен- Н4-фолата может превращаться в другие одноуглеродные группы (фрагменты): метенильную (-СН=), формильную (-НС=О), метильную (-СН3) и формиминогруппу (-CH=NH
В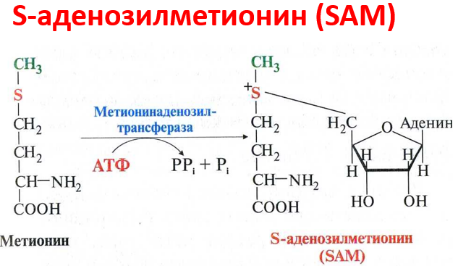 се
образующиеся производные Н4-фолата
играют роль промежуточных переносчиков
и служат донорами одноуглеродных
фрагментов при синтезе некоторых
соединений: пуриновых оснований и
тимидиловой кислоты (необходимых для
синтеза ДНК и РНК), регенерации метионина,
синтезе различных формиминопроизводных
(формиминоглицина и т.д.)
се
образующиеся производные Н4-фолата
играют роль промежуточных переносчиков
и служат донорами одноуглеродных
фрагментов при синтезе некоторых
соединений: пуриновых оснований и
тимидиловой кислоты (необходимых для
синтеза ДНК и РНК), регенерации метионина,
синтезе различных формиминопроизводных
(формиминоглицина и т.д.)
Обмен фенилаланина и тирозина. Пути превращения в норме.
Метаболизм фенилаланина
Основное количество фенилаланина расходуется по 2 путям:
• включается в белки;
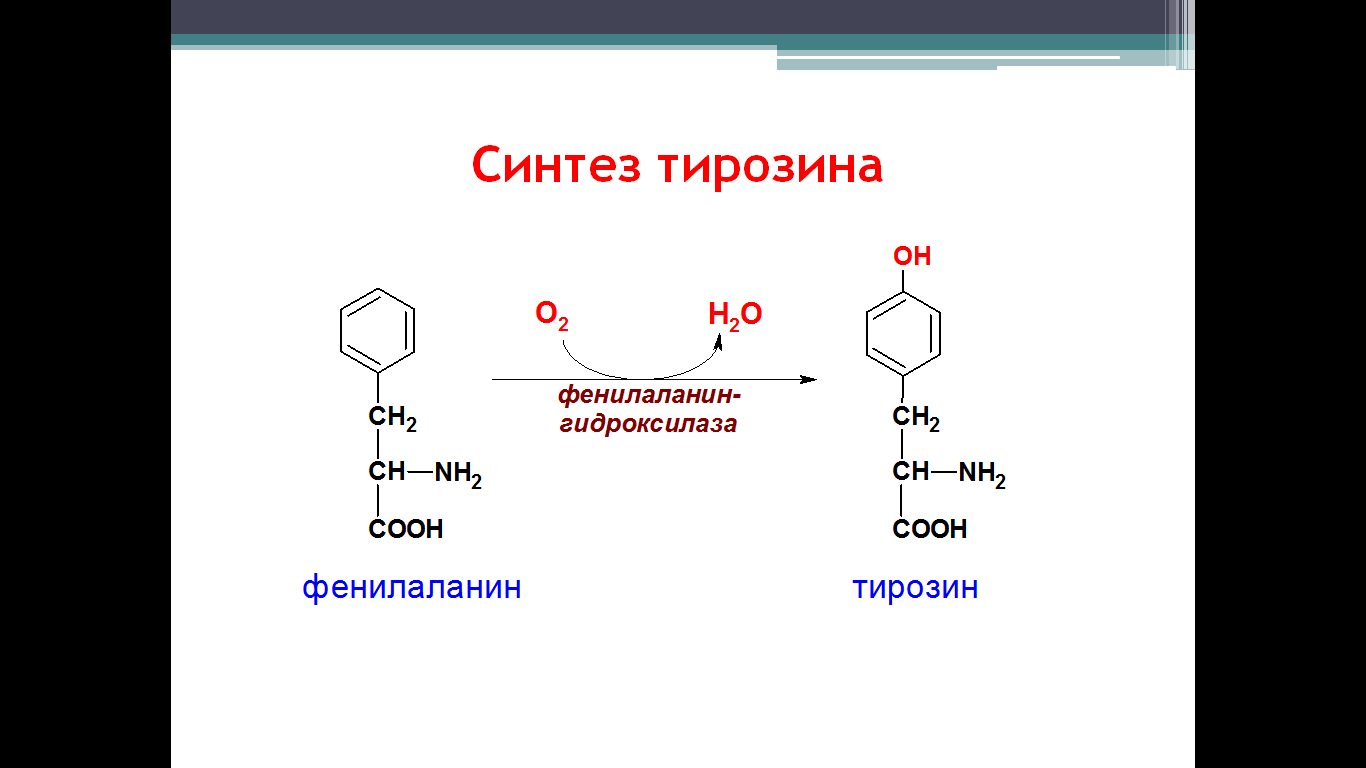
• превращается в тирозин. Превращение фенилаланина в тирозин прежде
всего
необходимо для удаления избытка
фени-лаланина, так как высокие концентрации
его токсичны для клеток. Образование
тирозина не имеет большого значения,
так как недостатка этой аминокислоты
в клетках практически не бывает.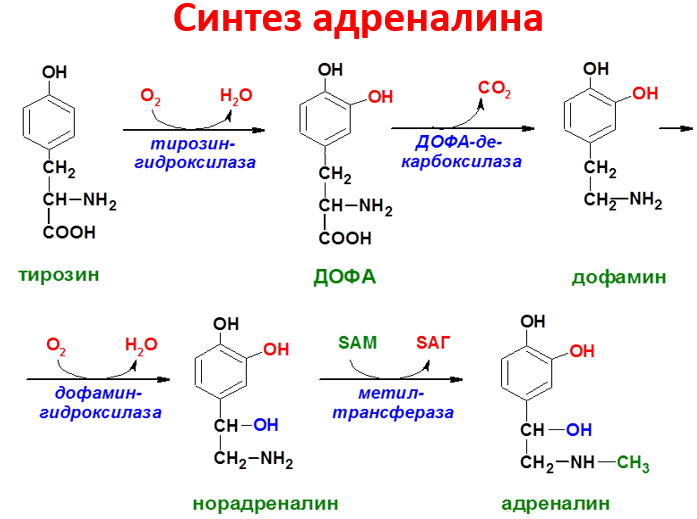
Фенилаланин - незаменимая аминокислота, так как в клетках животных не синтезируется её бензольное кольцо.
Тирозин - условно заменимая аминокислота, поскольку образуется из фенилаланина. Содержание этих аминокислот в пищевых белках (в том числе и растительных) достаточно велико. Фенилаланин и тирозин используются для синтеза многих биологически ктивных соединений. В разных тканях метаболизм этих аминокислот происходит по-разному
М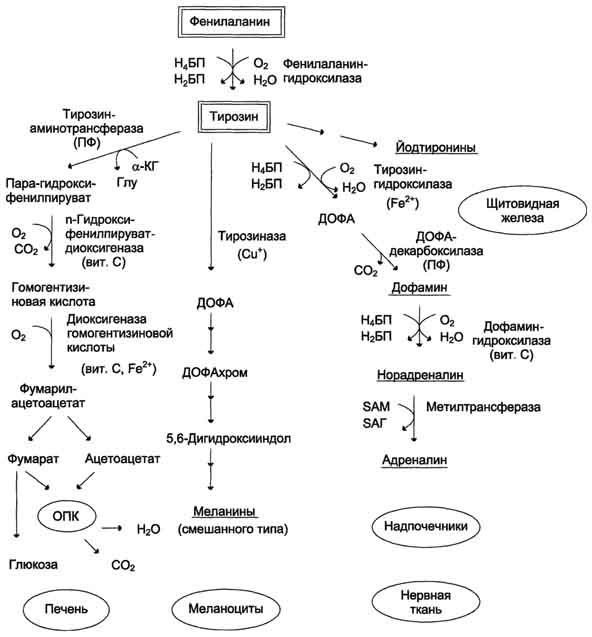 етаболизм
феиилаланина
етаболизм
феиилаланина
Основное количество фенилаланина расходуется по 2 путям:
включается в белки;
превращается в тирозин.
Превращение фенилаланина в тирозин прежде всего необходимо для удаления избытка фенилаланина, так как высокие концентрации его токсичны для клеток. Образование тирозина не имеет большого значения, так как недостатка этой аминокислоты в клетках практически не бывает. Основной путь метаболизма фенилаланина начинается с его гидроксилирования, в результате чего образуется тирозин. Эта реакция катализируется специфической монооксиге-назой - фенилаланингидр(жсилазой, кофермен-том которой служит тетрагидробиоптерин (Н4БП). Активность фермента зависит также от наличия Fe2+. Реакция необратима. Н4БП в результате реакции окисляется в дигидробиоптерин (Н2БП). Регенерация последнего происходит при участии дигидроптеридинредуктазы с использованием NADPH + H+. Обмен тирозина значительно сложнее, чем обмен фенилаланина. Кроме использования в синтезе белков, тирозин в разных тканях выступает предшественником таких соединений, как катехоламины, тироксин, меланины, и ка-таболизируется до СО2 и Н2О.
Катаболизм тирозина в печени
В печени происходит катаболизм тирозина до конечных продуктов. Специфический путь катаболизма включает несколько ферментативных реакций, завершающихся образованием фумарата и ацетоацетата Трансаминирование тирозина с ос-кетоглутаратом катализирует тирозинаминотрансфе-раза(кофермент ПФ) - индуцируемый фермент печени млекопитающих. В результате образуется п-гидроксифенилпируват.
В реакции окисления п-гидроксифенилпирувата в гомогентизиновую кислоту происходит декарбоксилирование, гидроксилирование ароматического кольца и миграция боковой цепи. Реакцию катализирует фермент n-гидроксифенилпируватдиоксигеназа, кофакторами которого выступают витамин С и Fe2+.
Превращение гомогентизиновой кислоты в фумарилацетоацетат сопровождается расщеплением ароматического кольца. Эта реакция катализируется диоксигеназой гомогентизиновой кислоты, в качестве кофермента содержащей Fe2+.
Обмен фенилаланина и тирозина связан со значительным количеством реакций гидроксилирования, которые катализируют оксигеназы. Ферменты оксигеназы (гидроксилазы) используют молекулу О2 и кофермент-донор водорода (чаще - Н4БП). Для катализа оксигеназам необходимы кофакторы - Fe2+ или гем (для некоторых - Сu+), а для многих ещё и витамин С. Оксигеназы делят на 2 группы:
Монооксигеназы - один атом О2 присоединяют к продукту реакции, другой используют для образования Н2О;
Диоксигеназы - оба атома О2 используют для образования продукта реакции.
Почти все процессы расщепления ароматических колец в биологических системах катализируются диоксигеназами, подклассом ферментов, открытым японским биохимиком Осами Хайяши. В результате разрыва бензольного кольца образуется малеилацетоацетат, который в процессе цис- и транс-изомеризации превращается в фумарилацетоацетат.
Гидролиз фумарилацетоацетата при действии фумарилацетоацетатгидролазы приводит к образованию фумарата и ацетоацетата. Фумарат может окисляться до СО2 и Н2О или использоваться для глюконеогенеза. Ацетоацетат - кетоновое тело, окисляемое до конечных продуктов с выделением энергии.
Превращение тирозина в меланоцитах. В пигментных клетках (меланоцитах) тирозин выступает предшественником тёмных пигментов - меланинов. Среди них преобладают 2 типа: эумеланины и феомеланины. Эумеланины (чёрного и коричневого цвета) - нерастворимые высокомолекулярные гетерополимеры 5,6-дигидроксииндола и некоторых его предшественников. Феомеланины - жёлтые или красновато-коричневые полимеры, растворимые в разбавленных щелочах. Находятся они, в основном, в составе волос. Меланины присутствуют в сетчатке глаз. Цвет кожи зависит от распределения меланоцитов и количества в них разных типов меланинов.