Материал: Хромосомы эукариот
Образование прицентромерного гетерохроматина является результатом модификации гистонов и прежде всего метилированием лизина в молекуле гистонов Н3 и Н4. Метилированные остаток лизина узнают различные белки, которые запускают процессы, связанные с «молчанием». Например, у инфузории с метилированным лизином связываются белки Pddq, Pdd3, что вызывает диминуцию хроматина. У млекопитающих и дрозофилы модифицированный гистон Н3 может взаимодействовать с белком НР1 (Heterochromatin protein 1), который в свою очередь может привлекать множество других белков. Полагают, что связь НР1 с модифицированным гистоном Н3 необходима для стабилизации репрессированного состояния гетерохроматина.
Инактивация Х-хромосомы млекопитающих сопровождается гипоацетилированием гистонов Н2А, Н3 и Н4 и метилированием гистона Н3.
Другим вариантом модификации гистонов является их фосфорилирование - дефосфорилирование, что обеспечивает декомпактизацию хроматина при активации транскрипции и конденсацию хромосом во время деления клетки или апоптоза. Баланс фосфатных групп поддерживают киназы и фосфотазы.
Образец модификаций молекул гистонов называют «гистоновым кодом». Гипотеза «гистонового кода» предполагает, что эти модификации являются предпосылкой для последующего изменения структуры хроматина и стабильно поддерживают характер экспрессии генов в ходе клеточных делений. Посттрансляционные модификации гистонов (ацетилирование, метилирование, фосфорилирование и др.) создают специфические сайты связывания для регуляторных белков хроматина.
Формирование гетерохроматина строго детерминировано и происходит на определенной стадии эмбрионального развития, когда начинается транскрипция на хромосомах развивающегося эмбриона. Формирование гетерохроматиновых доменов надежно защищает организм от экспрессии генетического материала, которая не нужна в соматических клетках.
Помимо образования гетерохроматина другим механизмом «выключения» некодирующих последовательностей является диминуция хроматина. Это программируемая элиминация части генетического материала из генома презумптивных соматических клеток.
У циклопов в 3-7 делениях дробления элиминируется от 10 до 94% генома germ line. Цитологическая картина диминуции хроматина видоспецифична. Доля элиминируемой ДНК не зависит от исходного размера генома. Например, 2 вида циклопов Cyclops kolensis, Cyclops insignis обитают в одинаковых условиях, количество ДНК в диплоидных клетках примерно одинаково - 4,6 пг и 4,3 пг. У C. kolensis после диминуции остается лишь 6% генома. Однако при этом количество хромосом не изменяется (22), хотя размеры их становятся существенно меньше. У C. insignis диминуции хроматина не происходит.
У циклопов перед диминуцией хроматина происходит удлинение интерфазы в 8-9 раз. В конце этой интерфазы появляются плотные гранулы хроматина (до 600). Далее они сливаются, вокруг них формируется плотная однослойная без пор мембрана, непрницаемая для факторов декомпактизации. Внутри этих гранул происходит лизис ДНК.
Генетический аппарат ресничных инфузорий отличается ядерным диморфизмом. Он проявляется в наличии в клетке двух типов ядер: микронуклеуса (Ми) и макронуклеуса (Ма). Ми служит генеративным ядром, используемым для передачи наследственной информации в ряду поколений. В вегетативно растущих клетках гены Ми не транскрибируются. Зато активно транскрибируются гены Ма, выполняющего роль рабочего ядра и контролирующего процессы жизнедеятельности.
При конъюгации у инфузорий две клетки прикладываются друг к другу и плавают вместе 10-12 часов, а затем расходятся. За время конъюгации их Ма начали разрушаться, а Ми поделились путем мейоза на четыре гаплоидных ядра каждое. Ход последующих событий варьирует в деталях у разных видов инфузорий, но принципиальная схема общая: клетки партнеров обмениваются гаплоидными ядрами, по одному от каждой клетки, затем каждое сливается с местным (стационарным) гаплоидным ядром, то есть происходит оплодотворение. К этому моменту все лишние ядра дегенерируют, и в каждой клетке остается по одному диплоидному ядру - продукту оплодотворения. После расхождения партнеров ядро делится митотически на два. Одно из дочерних ядер останется Ми, другое превратится в Ма. Развитие Ма занимает несколько дней и сопровождается полной реорганизацией генома Ми-предшественника.
ДНК хромосом Ми имеет большие размеры, характерные для эукариотических хромосом. Гены в них собраны в группы с длинными промежутками между ними, заполненными разнообразными уникальными и повторяющимися последовательностями ДНК. Важная особенность генов Ми заключается в том, что все они прерваны особыми элементами, называемым IES (internal eliminated sequences). Кодирующие участки генов, разделенные IES, называют MDS <#"870758.files/image004.gif">
Рисунок 4. Дифференцировка Ма у
брюхоресничных инфузорий: а - общая схема дифференцировки ядер из
оплодотворенного ядра; б - схема превращения микроядерного гена в ген Ма
Перестройка генетического материала происходит при развитии Ма из Ми. Она начинается сразу после конъюгации с множественных раундов репликации хромосом в исходном Ми без его деления. Это приводит к образованию политенных хромосом. Политенизация хромосом увеличивает их размеры.
Во время политенизации происходит важное для реорганизации генома Ми удаление всех IES. Оно осуществляется путем множественных событий рекомбинации по прямым повторам ДНК, имеющимся на концах каждого IES. У E. crassus IES несут концевые повторы от 2 до 4 п.н. и всегда содержат димер 5'-TA, у O. nova - от 2 до 19 п.н. и не содержат 5'-TA. Поскольку IES прерывают кодирующие участки генов (MDS), состыковка последних при вырезании IES (в виде колец) происходит с точностью до нуклеотида по той же схеме, что и аналогичная рекомбинация у бактерий. Помимо коротких IES ДНК Ми E. crassus содержит два родственных элемента размером около 5,3 т.п.н., называемых TEC1 и TEC2. Эти элементы принадлежат к подвижным генетическим элементам. TEC-элементы представлены в Ми примерно 30 тыс. копиями и все вырезаются в виде колец.
После политенизации и удаления IES и подвижных элементов происходит разрезание политенных хромосом на фрагменты, соответствующие отдельным генам. Механизм фрагментации хромосом пока невыяснен. У равноресничных инфузорий, к которым принадлежит всем известная инфузория-туфелька, обнаружены особые участки, в которых происходят разрывы хромосом. Их называют CBS <#"870758.files/image005.jpg">
Рисунок 5. Диминуция хроматина у
Parascaris univalens: а - цитологическая картина диминуции хроматина
На левой схеме изображены анафазы второго деления дробления на стадии двух клеток. Диминуция происходит в верхней клетке С1. В нижней клетке Г1 идет нормальный митоз. Справа представлена стадия четырех клеток после завершения второго деления дробления; б - схема дифференцировки клеток зародышевого пути и соматических клеток на ранних стадиях эмбрионального развития аскариды. Пресоматические клетки С1, С2, С3 и С4, подвергшиеся диминуции, изображены кружками, окруженными четырьмя точками.
У P. univalens при диминуции удаляется около 85% хроматина. Основная часть удаляемого материала состоит из сателлитной ДНК. Однако при диминуции удаляются и некоторые уникальные гены, например ген ALEP1, который кодирует определенный рибосомный белок. Этот белок синтезируется только в генеративных клетках и отсутствует в соматических. Таким образом, элиминируемый хроматин содержит не просто ненужную ДНК, но необходим для клеток зародышевого пути. Предполагается, что элиминация гена ALEP1 вызывает структурные различия между рибосомами зародышевых и соматических клеток, которые могут обусловливать дифференциальную трансляцию специфических мРНК в двух типах клеток.
Эффект диминуции хроматина указывает на то, что большая часть избыточной ДНК не несет ни кодирующих, ни регуляторных функций. Функции избыточной ДНК как-то связаны с клетками germ line. Возможно, что некодирующая часть генома играет важную роль в изоляции видов, осуществляемой в мейозе при конъюгации хромосом.
При случайном скрещивании видов с диминуцией хроматина и без диминуции: В яйцеклетке в ее цитоплазме у видов с диминуцией хроматина есть ферменты, участвующие в диминуции хроматина. В результате диминуции хроматина могут подвергнуться структурные гены отцовского происхождения. Потеря структурных генов - гибель гибридного зародыша.
При обратном направлении скрещивания отцовские хромосомы не пройдут через цикл диминуции хроматина, так как в яйцеклетке не будет нужных ферментов. В результате отцовские структурные гены останутся в неактивной форме. Зародыш тоже погибнет.
. Строение хромосом эукариот
Хромосома - это
самовоспроизводящийся структурный элемент ядра клетки. Число, размер и форма
хромосом строго определены и специфичны для каждого вида. Термин «хромосома»
был предложен в 1888 W. Waldeyer. Каждая хромосома состоит из одной или
нескольких пар хроматиновых нитей. Морфологию хромосом эукариот определяют на
стадии метафазы митоза. Характерные для вида особенности хромосом (то есть их количество,
размеры, форма, наличие спутников и т.д.) называют кариотипом.

Рисунок 6. Примеры кариотипов
Рисунок 7. Идиограмма хромосом
человека
В объеме интерфазного ядра каждое
плечо хромосомы располагается в определенной зоне, объем которой не перекрывает
объем соседних хромосом, хотя они примыкают друг к другу. Каждая из хромосом в
нескольких местах связана с ядерной оболочкой. Все теломерные участки
фиксированы на ядерной мембране на одном полюсе интерфазного ядра; на
противоположном полюсе ядра располагаются (тоже фиксированные на мембране)
центромерные районы хромосом. В последнее время показано, что в нормальных
клетках богатые генами хромосомы располагаются ближе к центру ядра, а бедные
генами хромосомы - на периферии ядра. Например, хромосом 18 и 19. Материал
хромосомы 19, богатой генами, тяготеет к внутреннему компартменту ядра, тогда
как территория хромосомы 18, бедной генами, находится на его периферии.
Область, прилежащая к ядерной оболочке, и области, прилежащие к конститутивному
(н-р, прицентромерному) гетерохроматину, являются в ядре неактивными
компартментами. Простое перемещение генов в эти компартменты приводит к их
долговременной инактивации. Пример: в эритроидных клетках глобиновые гены
располагаются далеко от конститутивного гетерохроматина, тогда как в
неэритроидных клетках они практически контактируют с конститутивным
гетерохроматином.
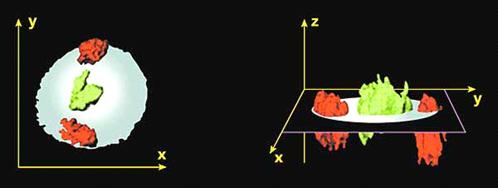
Рисунок 8. Пространственное
расположение хромосом в объеме ядра
Каждая хромосома имеет первичную перетяжку или центромеру, которая делит хромосому на два плеча. Участок центромеры ответственен за контакт с микротрубочками веретена деления и перемещения хромосомы при клеточном делении. К центромерной ДНК присоединяются центромерные белки, образующие кинетохор. Это справедливо для хромосом с локализованной центромерой.
У некоторых растений, грибов и
членистоногих (скорпионы) хромосомы имеют диффузную центромеру. Это означает,
что контакт с микротрубочками веретена осуществляется по всей длине хромосомы.
Такие хромосомы называют голоцентрическими.
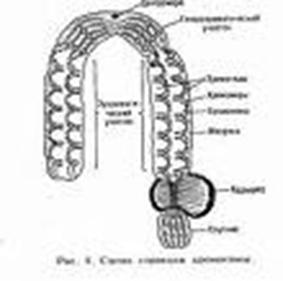
Рисунок 9. Строение хромосом
Описан и третий тип хромосом у некоторых видов аскарид. В эмбриогенезе у Ascaris megalocephala в первых двух делениях дробления выявляются крупные хромосомы с нормально функционирующей локализованной центромерой. Однако при третьем делении в трёх бластомерах из четырёх хромосомы распадаются на мелкие фрагменты, каждый из которых ведёт себя как обычная хромосома; т.е. расщепляется на хроматиды, расходящиеся к полюсам. Часть хромосомного материала при этом разрушается. Каждая из мелких хромосом обладает собственной центромерой. Таким образом, исходные крупные хромосомы аскариды, сохраняющиеся в четвёртом бластомере, из которого развиваются клетки зародышевого пути, являются потенциально полицентрическими хромосомами. Однако, в первых двух митозах дробления и в клетках зародышевого пути эти многочисленные центромеры не активны.
В центромерных районах хромосом локализована сателлитная ДНК, т.е. высоко повторяющиеся последовательности ДНК. Центромерная ДНК S. cerevisiae состоит из повторяющихся участков по 110 п.н. Она имеет по 2 консервативных участка (1 и 3) и центральный элемент (2), обогащённый АТ. У мыши - основной класс стДНК - повтор последовательности в 234 п.н.
ДНК центромерного участка хромосом человека представлена блоком тандемно организованных повторяющихся единиц размером 171 пн. Тяжи мономеров включают до 3 млн. пн. В середине тяжа идентичность копий мономеров составляет 99%, а на концах тяжа единицы мономеров более дивергированы. Характер организации центромер человека и арабидопсиса сходен, хотя первичные последовательности их мономеров совершенно различны.
Центромера Х-хромосомы дрозофилы состоит в основном из двух типов очень коротких простых сателлитов, ААТАТ и ААГАГ, прерываемых ретротранспозонами.
В центромерном хроматине эукариот обнаружены видоспецифические варианты гистона Н3 - CENP-A у человека и CENH3 у растений. Предполагается, что именно CENH3, взаимодействуя другими гистонами Н2А, Н2В, Н4 формирует и определяет специфический тип нуклеосом, которые присутствуют только в функционирующих центромерах. Возможно, что такие нуклеосомы являются «якорями» для образования кинетохора.
В организации центромерного хроматина определенную роль могут играть малые РНК. Показано, что пул специфических для центромер малых РНК (40-200 пн) транскрибируется с центромерного ретротранспозона и сателлитной ДНК, связан с центромерным гистоном Н3 в пределах комплекса кинетохора кукурузы.
Уникальные данные получены о центромере риса. Центромеры хромосом риса содержат сателлитные повторы с размером мономера 155 пн. В хромосоме 8 длина этого тяжа наименьшая среди 12 хромосом риса - около 64 тпн. Общая длина центромерного участка хромосомы 8 составляет 2 млн. пн. Внутри этого района есть участок непосредственного формирования кинетохора (около 750 тпн). Но самое интересное - в составе центромеры кроме ретротранспозонов обнаружены активно транскрибируемые гены, 48 из которых имеют последовательности высоко гомологичные известным белкам или кДНК-клонам. Более того, 14 предполагаемых генов обнаружены в области кинетохора. Это опровергает мнение, что центромеры не содержат активно транскрибируемых генов.
Сформированный кинетохор представляет собой трехслойную структуру: внутренний слой (40-60нм), светлоокрашенный средний слой (25-30 нм) и внешний слой (40-60 нм). Внутренняя поверхность кинетохора взаимодействует с центромерным хроматином. Микротрубочки, связанные с кинетохором, контактируют с его внешним слоем.
В зависимости от расположения центромеры выделяют метацентрические, субметацентрические и акроцентрические хромосомы. Для акроцентрических хромосом характерно наличие вторичной перетяжки, отделяющей спутники. Данный участок хромосомы содержит большое количество генов рРНК. Гены рРНК часто образуют тандемные пары, которые организованы в кластеры. Каждый кластер тандемно повторяющихся генов рРНК соответствует ядрышковому организатору.