Материал: Жукова Л.А. (отв. ред.) Онтогенетический атлас лекарственных растений. Том VII
ство цветков на особь (до 83,2 ± 4,2 шт.), начинается формирование плодов. Наблюдается также усложнение соцветий: от кисти к раскидистой метелке. Продолжительность средневозрастного генеративного состояния составляет 14 ± 1 суток.
В СТАРОЕ ГЕНЕРАТИВНОЕ состояние растения переходят в первой декаде августа. В этом возрастном состоянии начинаются некротические процессы в главном корне, постепенно в прикорневой розетке накапливаются отмершие засохшие листья, созревают плоды. Высота генеративных побегов и соцветий достигает максимальных значений (55,6 и 39,8 см соответственно). Количество цветков на особь уменьшается до 52,6 ± 2,6 шт. Общий период цветения растений варьирует от 42 до 54 суток. Продолжительность данного возрастного состояния составляет 36 ± 2 суток. Вегетация заканчивается в третьей декаде сентября. После плодоношения растение отмирает.
Таблица 6
Морфометрические показатели генеративных органов колокольчика сибирского в разных онтогенетических состояниях
|
- |
Высота, см |
Количество на особь, шт. |
||
|
Состоя ние |
|
|
|
|
Вид |
генеративного |
соцветия |
генеративных |
цветков |
|
|
|
побега |
|
побегов |
|
|
g1 |
38,6 ± 1,9 |
17,3 ± 0,8 |
1,0 ± 0,0 |
18,1 ± 0,9 |
С. sibirica |
|
|
|
|
|
g2 |
46,1 ± 2,3 |
21,6 ± 1,2 |
1,0 ± 0,0 |
83,2 ± 4,2 |
|
|
|
|
|
|
|
|
g3 |
55,6 ± 2,9 |
39,8 ± 1,9 |
1,0 ± 0,0 |
52,6 ± 2,6 |
|
|
|
|
|
|
Рис. 19. Онтогенез колокольчика сибирского (Campanula sibirica L.)
Таким образом, онтогенез С. sibiricа длится два вегетационных сезона и проходит по А-типу (Жукова, 1995). Постгенеративный период у данного вида не выражен.
131
13.Онтогенез торилиса японского
(Torilis japonica (Houtt.) DC.)
Torilis japonica (Houtt.) DC. (сем. Apiaceae) является типичным представителем малолетних монокарпических растений с пластичным временным переходом к репродукции. С учетом классификации И. Г. Серебрякова (1962, 1964) его можно охарактеризовать как стержнекорневой длительно вегетирующий яровой или озимый однолетник или двулетник с удлиненными или полурозеточными прямостоячими побегами.
Главный корень тонкий (2–4 мм в диаметре) веретеновидный, разветвленный до III порядка. Цветоносный побег высотой 30(11)–130 см, полурозеточный или удлиненный, почти от основания ветвящийся до IV–V порядков. Стебель тонкоребристый, опушенный короткими, направленными вниз волосками, диаметром 0,3–0,7 см. До терминального двойного зонтика развивается 6–7 стеблевых листьев. Нижние – дважды или трижды перистосложные, в очертании продолговато-яйцевидные или треугольные, длиной 7,5–8 см, шириной 4,5–5 см. Листочки первого порядка яйцевидные, второго – продолговато-ланцетные перистораздельные, конечные элементы ланцетовидные, на верхушке оттянутые. Жилкование перисто-сетчатое. Черешок длиной до 1,4 см, желобчатый. Влагалище длиной 1,3–1,5 см, узкое, с белопленчатым краем. Все части листа (листовая пластинка преимущественно по жилкам) покрыты жесткими белыми прижатыми волосками. Самые верхние листья менее расчлененные, нередко тройчатосложные, без черешков.
Соцветие – метелка из двойных зонтиков. Терминальный двойной зонтик около 2–5 см в диаметре, состоит из 5–12 неравных по длине, покрытых щетинистыми волосками лучей. Обертка и оберточка – из 4– 10 линейных щетинисто-волосистых листочков. Простые открытые зонтички включают до 14 цветков, из них краевые – крупные, слабо зигоморфные. Цветки обоеполые или мужские. Чашелистики треугольноланцетные, длиной около 1 мм, опушенные. Лепестки белые или розоватые, с загнутой внутрь верхушкой, длиной 2 мм, на абаксиальной стороне опушенные. Пыльники розово-фиолетовые. Подстолбия коротко конические, стилодии расходящиеся или отогнутые на спинную сторону мерикарпиев. Плоды в очертании яйцевидные или эллиптические, длиной 2–4 мм, густо покрыты щитинками и шипами, карпофор наполовину двураздельный. Мерикарпии с первичными и вторичными слабо выступающими ребрами, покрытыми разной длины щетинками.
На территории России T. japonica встречается в европейской ее части, в Предкавказье и на Дальнем Востоке. Общее распространение:
132
Средняя и Атлантическая Европа, Средиземноморье, Кавказ, Малая Азия, Иран; в Северной Америке, Японии и Китае – как заносное (Шишкин, 1950; Флора Вост. Европы,2004).
Вид произрастает в разреженных широколиственных, хвойно-широ- колиственных, сосновых и пихтовых лесах, осинниках, березняках, ольшаниках, среди кустарников, на лесных полянах и опушках, у дорог, в населенных пунктах (Зонтичные Средней России, 1997; Пименов, Остроумова, 2012).
Наблюдения за индивидуальным развитием растений и сбор материала проводили в г. Пущино Московской области в смешанном лесу на окраине города (Петрова, 2008), а также близ учебно-научного центра ВГУ «Веневитиново» (Воронежская область).
Периодизация онтогенеза торилиса японского проведена в соответствии с классическими работами Т. А. Работнова (1950) и А. А. Уранова (1974), а также с учетом современных данных (Онтогенетический атлас растений, 2007; 2011). Онтогенез представлен на рисунке 20.
СЕМЕНА небольших размеров (2,5–3,5 мм). Вес 1 000 семянок составляет 1,5–2 г. (Рысина, 1973). Зародыш длиной около 0,30–0,42 мм, состоит из двух семядолей, приблизительно равной им по длине осевой части (гипокотиль, зародышевый корень) и верхушечной почечки, представленной группой меристематических клеток.
ПРОРОСТКИ. Прорастание начинается или осенью (почти сразу после опадения с материнского растения), или в конце весны и часто растягивается до середины июля. По данным Г. П. Рысиной (1973), в Подмосковье массовое прорастание семян наблюдается во второй половине апреля. Согласно исследованиям американских ученых (Baskin J. & Baskin C., 1975), при прорастании T. japonica в поле (штат Кентукки) и в лабораторных условиях установлено, что отрицательные зимние температуры обычно вызывают состояние покоя семян торилиса японского, которое снимается лишь весенне-летним режимом температур и увлажнения. В связи с этим наиболее активное образование всходов происходит либо осенью в год плодоношения, либо начиная с августа следующего вегетационного сезона, что ведет к развитию растений по озимому типу.
Семядоли с продолговато-ланцетной, длиной 10–18 мм и шириной 1–2 мм, гладкой, на верхушке с белым острием пластинкой, плавно переходящей в приблизительно равный ей по длине черешок. Жилкование перистое, боковые жилки слабо выражены. Гипокотиль длиной 10– 17 мм, часто имеет фиолетовую окраску. Главный корень короткий, с непротяженной зоной корневых волосков, заглубляется в почву до
12 мм.
133
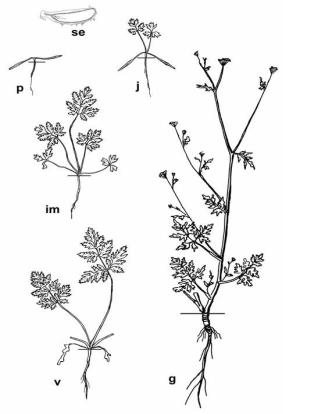
Рис. 20. Онтогенез торилиса японского (Torilis japonica (Houtt.) DC.)
ЮВЕНИЛЬНЫЕ растения имеют розеточный побег. Пластинка первого листа трехрассеченная, в очертании широко-яйцевидная, длиной 7–14 мм, шириной 12–20 мм, опушена немногочисленными белыми жесткими шиловидными волосками, поверхность которых покрыта округлыми протуберанцами. Центральный сегмент трехраздельный, боковые неравно двух-трехраздельные или лопастные. Черешок желобчатый, длиной 18–31 мм. Влагалище узкое. Второй лист длинночерешковый, тройчатосложный. Боковые листочки перистораздельные или рассеченные, в очертании яйцевидные; сегменты их по краю надрезанные. Центральный листочек трехрассеченный, ромбовидный, с трехраздельными сегментами. Третий лист дваждыперистосложный, с двумя парами листоч-
134
ков первого порядка. Последующие листья напоминают структуру третьего, однако увеличивается количество пар листочков I и II порядков. Жилкование перисто-краебежное. Все части листьев опушены редкими белыми шиловидными волосками. Главный корень ювенильного растения тонкий, длиной 7–8 см, слабо ветвящийся (не выше II порядка).
ИММАТУРНЫЕ растения сохраняют розеточный тип побега, на котором наряду с 2–3 засыхающими ювенильными листьями имеется до 3–4 дваждыперистосложных листов с 3 парами боковых листочков. Семядоли отмирают. Главный корень удлиняется, однако интенсивность ветвления при этом не увеличивается.
ВИРГИНИЛЬНЫЕ растения. У одних особей, обычно развивающихся по двулетнему циклу, побег сохраняет розеточность на протяжении всего прегенеративного периода. Такие растения уходят под снег с несколькими розеточными листьями, имеющими пазушные почки, и верхушечной почкой, прикрытой влагалищами редуцированных листьев. У других особей, онтогенез которых идет по типу яровых однолетников, побег становится полурозеточным за счет образования новых метамеров с удлиненными междоузлиями. Вскоре они переходят к репродукции. Особи с обоими типами развития встречаются в одной популяции, что, вероятно, уменьшает внутри- и межвидовую конкуренцию.
Гипокотиль виргинильных растений слегка разрастается и по всей длине опробковевает, из-за чего цвет его становится коричневым. Главный корень начинает интенсивно ветвиться, при основании утолщается; у озимых и двулетних форм в результате контрактильной деятельности он приобретает слабую поперечную морщинистость.
ГЕНЕРАТИВНЫЕ растения. Отметим, что четких критериев для выделения генеративного периода торилиса японского выявить не удалось.
Яровые однолетники переходят к цветению в год прорастания, цветение начинается в июле – начале августа, плодоношение растягивается до конца сентября. Озимые и двулетние формы зацветают во втором вегетационном сезоне, в июне – начале июля, а плодоносят – в августе. Таким образом, T. japonica присущ неустойчивый средне-позднелетний ритм цветения. В гербарии нами был обнаружен интересный экземпляр цветущей особи, собранный в Касимовском районе Рязанской области 1 августа на опушке смешанного леса: растение в высоту достигало всего 11 см и до соцветия имело только 5 листьев ювенильного типа.
Для яровых форм характерен полурозеточный прямостоячий мезатонно ветвящийся моноциклический побег. Как правило, в пазухах нескольких нижних междоузлий формируются слабо развитые паракла-
135